Key words
pharmacology - toxicology - apoptosis
Introduction
The thiazolidinedione (TZDs) are used in patients with diabetes to help improve insulin
resistance and glucose homeostasis. Furthermore, these drugs are known as peroxisome
proliferator-activated receptor gamma (PPARγ) agonists [1]
[2]
[3]
[4]. These synthetic compounds are pioglitazone, rosiglitazone, troglitazone and ciglitazone
[5]
[6]. There are controversies in the relationship between the use of TZDs drugs, such
as pioglitazone (PG), and the cardiovascular outcomes. Some research has shown that
there is a positive relationship between the use of PG and heart failure [7]
[8]
[9]. In recent years, PG has been used to improve some brain disorders. However, little
is known about its neurotoxicity [10]
[11]
[12].
Many compounds have caused mitochondrial toxicity and these organelles has become
one of the research targets [13]
[14]. Brain and heart tissues depend on mitochondria for their high energy consumption
and maintenance of their normal function [15]
[16]. Research has shown that mitochondria are one of the targets of PPARγ agonists [3]
[17]
[18]. Mitochondrial dysfunction has been reported by exposure to TZDs. Research shows
that TZDs increases the generation of reactive oxygen species (ROS) through disruption
of respiratory chain complexes I and III in mitochondria, collapse in mitochondria
membrane potential (MMP), cytochrome c release, mitochondrial swelling, and apoptosis
[5]
[19]
[20]
[21]
[22]
[23]. One of the mechanisms by which TZDs cause cytotoxicity is through the generation
of ROS. It has been reported that there is a direct relevance between the level of
ROS and the degree of cytotoxicity induced by these compounds [2]
[3]
[14]. Increased ROS generation induced by some TZDs lead to oxidation of vital components
within mitochondria (such as mitochondrial DNA, mtDNA) and also induction of apoptotic
signaling [21]. Incubation of isolated brain mitochondria with pioglitazone resulted in impairment
of complex III in the mitochondrial respiratory chain [17].
The mitochondrial organelle is considered the source and target of reactive oxygen
species (ROS). The electron transport chain in mitochondria is one of the most important
sources of ROS generation [24]
[25]
[26]
[27]. Research has shown that mitochondrial dysfunction has been implicated in the etiology
of many diseases [22]. The consequences of mitochondrial inhibition or disruption include the generation
of ROS, ATP depletion, and eventually cell death (apoptosis/necrosis). Mitochondria
as one of the important organelles in eukaryotic cells are involved in important physiological
processes including the generation of free radicals, energy production and cell death
[2]
[3]
[14]
[19]
[22]. Therefore, mitochondrial dysfunction can be dangerous for different cells and organs
due to insufficient ATP generation and excessive level of ROS [18]. The in vitro cytotoxicity investigations can be helpful in providing mechanistic
information, and this information can be useful in understanding the more detailed
clinical observations [19].
The effects of PG on mitochondria have not been fully studied. Therefore, we studied
the response of mitochondria isolated from the rat brain and heart to several concentrations
of PG by measuring succinate dehydrogenase (SDH) activity, ROS generation, MMP collapse,
mitochondria swelling and cytochrome c release.
Materials and Methods
Animals
Male Wistar rats (n=10), weighing 250–300 g were housed under standard conditions
(temperature 20–25°C, humidity 50–60%, 12 h light–dark cycle and free access to food
and water). The experimental protocols were approved by the Animal Ethics Committee
of Shahid Beheshti University of Medical Sciences. All efforts were made to minimize
the number and the suffering of animals used.
Mitochondria Isolation
In this study, mitochondria were isolated from the fresh brain and heart using a mitochondrial
isolation kit from Sigma Chemical Co. (St. Louis, MO, USA) according to the manufacture’s
instruction. The protein concentration of the pellet mitochondria was measured using
Bradford protein assay [28]. Furthermore, mitochondrial function was assessed through determining mitochondrial
SDH activity, mitochondrial reactive oxygen species (ROS) level, mitochondrial membrane
potential (MMP) collapse, mitochondrial swelling and cytochrome c release. In this
study, the mitochondrial purity and integrity were performed through the MTT test
(for evaluation of mitochondrial function/ mitochondrial complex II) and cytochrome
c oxidase (complex IV) assay kit, respectively.
Succinate Dehydrogenase (SDH) assay
Briefly, MTT dye was used to evaluate SDH activity. At first, mitochondria isolated
from the brain and heart were exposed to PG concentrations (12.5, 25 and 50 µg/ml)
for 30 min. Then, MTT (0.4%) was added to the mitochondrial suspension and incubated
at 37 °C for 30 min. In the final step, dimethyl sulfoxide (DMSO, 100 μl) were used
to dissolve formazan crystals, then the absorbance (570 nm) was assayed using an ELISA
reader (Tecan, Rainbow Thermo, Austria) [29].
ROS determination assay
2,7-dichlorofluorescein diacetate (DCFH-DA) probe at final concentration of 10 µM
was used to evaluate mitochondrial ROS generation. The isolated mitochondria from
brain and heart were suspended in respiration assay buffer and then were exposed to
PG concentrations (12.5, 25 and 50 µg/ml). Then, DCFH-DA was added to the mitochondrial
suspension and incubated for 5, 30 and 60 min at 37 °C. The fluorescence intensity
(EXλ= 488 nm and EMλ=527 nm) was assayed using a fluorescence spectrophotometer (Shimadzu
RF5000U) [30].
MMP determination assay
The Rhodamine 123 (Rh 123) probe at final concentration of 10 µM was used to evaluate
MMP collapse. The isolated mitochondria from brain and heart were suspended in MMP
assay buffer and then were exposed to PG concentrations (12.5, 25 and 50 µg/ml). Then,
Rh 123 was added to the mitochondrial suspension and incubated for 5, 30 and 60 min
at 37 °C. The fluorescence intensity (EXλ= 490 nm and EMλ=530 nm) was assayed using
a fluorescence spectrophotometer (Shimadzu RF5000U) [31].
Mitochondrial swelling
The isolated mitochondria from brain and heart were suspended in mitochondrial swelling
assay buffer and then were exposed to PG concentrations (12.5, 25 and 50 µg/ml). Then,
mitochondrial swelling was evaluated at 5, 30 and 60 min at 37 °C. The absorbance
(540 nm) was assayed using using an ELISA reader (Tecan, Rainbow Thermo, Austria)
[30].
Cytochrome c release
Briefly, cytochrome c release was evaluated using the Quantikine Rat/Mouse Cytochrome
c Immunoassay kit provided by R & D Systems, Inc. (Minneapolis, Minn.). The micro-plate
was used to pre-coating the monoclonal antibody specific for rat/mouse cytochrome
c. In the next step, conjugate (75 µL), standard and positive control (50 µL) were
added to each well of the micro-plate. Then, 1 µg of protein from each supernatant
fraction was added to the sample wells. All controls and standards, controls and samples
were added to the micro-plate (two wells), and then substrate solution (100 µl) was
added to micro-plate. Finally, stop solution (100 µl) was added to each well of micro-plate
and optical density was evaluated at 540 nm.
Statistical analysis
Results are presented as mean±SD. All statistical analyses were performed using GraphPad
Prism (version 5). The assays were performed 3 times. Statistical significance was
determined using the one-way ANOVA test, followed by the post hoc Tukey test. The
one-way ANOVA test was used as a specific statistical analysis for the determinations
of SDH activity, and cytochrome c release. In some experiments, the two-way ANOVA
test, followed by the post hoc Bonferroni test was also performed. The two-way ANOVA
test was used for the determinations of mitochondrial ROS level, MMP and mitochondrial
swelling. Statistical significance was set at P<0.05.
Results
PG decreased the SDH activity
The results showed that exposure to PG (12.5, 25 and 50 µg/ml) decreased SDH activity
in mitochondria isolated from the brain and heart ([Fig. 1]). Also, this decrease in SDH activity in mitochondria isolated was in a concentration-dependent
pattern (50>25>12.5 µg/ml). In fact, the decrease in absorbance indicates a decrease
in SDH activity.
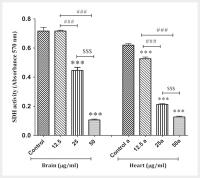 Fig. 1 SDH activity assay. The effect of PG on the SDH activity in the mitochondria isolated
from the brain and heart. Data are presented as mean±SD (n=3). The one-way ANOVA test
was carried out. *** show a significant difference in comparison with the corresponding control group
(P<0.001). ### Show a significant difference between 12.5 µg/ml group 25 and 50 µg/ml (P<0.001)
groups. $$$ Show a significant difference between 25 µg/ml group 50 µg/ml group (P<0.001).
Fig. 1 SDH activity assay. The effect of PG on the SDH activity in the mitochondria isolated
from the brain and heart. Data are presented as mean±SD (n=3). The one-way ANOVA test
was carried out. *** show a significant difference in comparison with the corresponding control group
(P<0.001). ### Show a significant difference between 12.5 µg/ml group 25 and 50 µg/ml (P<0.001)
groups. $$$ Show a significant difference between 25 µg/ml group 50 µg/ml group (P<0.001).
PG increased the ROS generation
In [Fig. 2], exposure of mitochondria isolated from the brain ([Fig. 2a]) and heart ([Fig. 2b]) to PG at all applied concentrations (12.5, 25 and 50 µg/ml) showed an increase
in the level of ROS generation. PG in a dose- and concentration-dependent manner increased
the level of ROS generation in isolated the brain and heart mitochondria.
 Fig. 2 ROS generation assay. The effect of PG on the ROS generation in the mitochondria
isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. ** and **** show a significant difference in comparison with the corresponding control group
(P<0.01 and P<0.0001, respectively).
Fig. 2 ROS generation assay. The effect of PG on the ROS generation in the mitochondria
isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. ** and **** show a significant difference in comparison with the corresponding control group
(P<0.01 and P<0.0001, respectively).
PG increased the MMP collapse
The results in [Fig. 3a] show that PG was able to collapse on MMP at concentrations of 25 and 50 µg/ml in
the isolated mitochondria from the brain and at concentration of 12.5 µg/ml had no
effect on MMP. However, PG at all concentrations caused the collapse of the MMP in
mitochondria isolated from the heart.
 Fig. 3 MMP collapse assay. The effect of PG on the MMP collapse in the mitochondria isolated
from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. **** show a significant difference in comparison with the corresponding control group
(P<0.0001).
Fig. 3 MMP collapse assay. The effect of PG on the MMP collapse in the mitochondria isolated
from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. **** show a significant difference in comparison with the corresponding control group
(P<0.0001).
PG increased the mitochondrial swelling
The results showed that exposure to PG (12.5, 25 and 50 µg/ml) at 5, 30 and 60 min
increased mitochondrial swelling in mitochondria isolated from the brain ([Fig. 4a]) and heart ([Fig. 4b]). Also, this increase in mitochondrial swelling in mitochondria isolated was in
a concentration-dependent pattern (50>25>12.5 µg/ml). In fact, the decrease in absorbance
indicates an increase in mitochondrial swelling.
 Fig. 4 Mitochondrial swelling assay. The effect of PG on the mitochondrial swelling in the
mitochondria isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. **** show a significant difference in comparison with the corresponding control group
(P<0.0001).
Fig. 4 Mitochondrial swelling assay. The effect of PG on the mitochondrial swelling in the
mitochondria isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The two-way ANOVA test was carried out. **** show a significant difference in comparison with the corresponding control group
(P<0.0001).
PG increased the cytochrome c release
In [Fig. 5], exposure of mitochondria isolated from the brain ([Fig. 5a]) and heart ([Fig. 5b]) to PG at all applied concentrations (12.5, 25 and 50 µg/ml) showed an increase
in the release of cytochrome c.
 Fig. 5 Cytochrome c release assay. The effect of PG on the cytochrome c release assay in
the mitochondria isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The one-way ANOVA test was carried out. * and *** show a significant difference in comparison with the corresponding control group
(P<0.05 and P<0.001). # , ## and ### show a significant difference between the control plus 25 µg/ml with 25 µg/ml plus
BHT and 25 µg/ml plus CsA (P<0.05, P<0.01 and P<0.001).
Fig. 5 Cytochrome c release assay. The effect of PG on the cytochrome c release assay in
the mitochondria isolated from the brain a and heart b. Data are presented as mean±SD (n=3). The one-way ANOVA test was carried out. * and *** show a significant difference in comparison with the corresponding control group
(P<0.05 and P<0.001). # , ## and ### show a significant difference between the control plus 25 µg/ml with 25 µg/ml plus
BHT and 25 µg/ml plus CsA (P<0.05, P<0.01 and P<0.001).
Considerably, the pretreatment of mitochondria with the MPT inhibitor (cyclosporine
A; Cs A) and an antioxidant (butylated hydroxytoluene; BHT), inhibited cytochromec
release from PG (25 µg/ml) treated mitochondria. Our results showed that PG release
of cytochrome c due to oxidative stress and MPT pore opening.
Discussion
Today, PG is used in the treatment of hyperglycemia in diabetic patients (type-2)
[10]
[32]. Research has shown that some TZDs induce mitochondrial dysfunction through different
mechanisms, including increase in ROS level, collapse in the MMP, mitochondrial swelling,
and induction of apoptosis signaling [5]
[19]
[20]
[21]
[22]
[23]. However, mechanistic information regarding exposure to PG and mitochondrial dysfunction
is not available. In this study, we investigated mitochondrial function after exposure
to different concentrations of PG. Furthermore, the functions of freshly mitochondria
(mitochondria isolated from the rat brain and heart) were assessed by measuring SDH
activity, ROS generation, MMP collapse, mitochondrial swelling, and cytochrome c release.
The brain tissue holds nearly 2% of total body mass, but consumes nearly 20% of total
body energy (ATP). In fact, the brain is one of the tissues that needs a lot of ATP.
Mitochondrion is known as a source of energy in the body, and it produces energy through
the respiratory chain. Therefore, brain tissue needs mitochondria to maintain its
normal function and energy consumption. Mitochondrial dysfunction in the brain is
associated with neurodegenerative diseases [15]
[33]
[34]. In addition, energy consumption in the heart is similar to that of the brain. On
the other hand, the heart needs energy for its normal function and development, and
mitochondria are the source of this energy [16]
[35]
[36].
In the mitochondria isolated from the brain and heart, we found a significant decrease
in SDH activity compared with control group following the addition of several concentrations
of PG. ROS are involved in important physiological processes including cell growth
and proliferation and apoptosis. Studies have shown that mitochondria are a major
source of ROS. The generation of ROS is via the electron leakage in the respiratory
chain of mitochondria, especially complexes I and III [24]
[37]
[38]
[39]. High levels of oxygen consumption can lead to increase generation of ROS in tissues
with high oxygen consumption (such as brain and heart). High levels of free radicals
lead to consequences including oxidative stress, damage to the mitochondrial membrane
and mtDNA, and induction of apoptosis [40]
[41]
[42]. Our findings regarding ROS generation using DCFH-DA shows that PG increases the
ROS levels in the mitochondria obtained from the brain and heart at 5, 30 and 60 min
after exposure. The results of this study are in agreement with other studies that
have shown that exposure to some TZDs increases the generation of ROS [3]
[14].
An increase in the level of ROS can induce the opening of mitochondrial permeability
transition (MPT) pore [2]. Research has shown that the opening of the MPT pore in the inner membrane leads
to the collapse of mitochondrial membrane potential, mitochondrial swelling, cytochrome
c release and subsequently induction of cell death (apoptosis) [4]
[13]. The MMP as one of the most important indicators of mitochondrial function can be
evaluated by fluorescence probes. Compared with the control group, exposure the mitochondria
obtained from the brain and heart with PG induced significant collapse in MMP. These
results are in agreement with the results of previous studies [2]
[14]
[20]
[21]. The collapse in MMP facilitates cytochrome c exit from mitochondria and induces
cell death [43].
Finally, the results showed that exposure of mitochondria isolated from the brain
and heart to PG caused mitochondrial swelling and cytochrome c release. Cytochrome
c release from mitochondria is one of the early events in the cell death process [43]. In conclusion, the results of this study suggest that pioglitazone increases the
generation of ROS through the effect on the mitochondrial respiratory chain. An increase
in the level of ROS can induce the opening of MPT pore. Finally, the opening of MPT
pore can disrupt the mitochondrial membrane, mitochondrial swelling, and cytochrome
c release and eventually cell death in the mitochondria isolated from the brain and
heart. Mitochondrial dysfunction in brain and heart is associated with neurodegenerative
and cardiovascular diseases.





