Key words
Curcumin -
Curcuma longa
- Zingiberaceae - lithium-pilocarpine model - [
18F]FDG PET - neuroinflammation
Abbreviations
[18F]FDG:
2-deoxy-2-[18F]Fluoro-D-Glucose
BW:
Body Weight
CT:
Computed Tomography
FITC:
Fluorescein Isothiocyanate
GFAP:
Glial Fibrillary Acidic Protein
MRI:
Magnetic Resonance Imaging
O. D.:
Optical Density
PET:
Positron Emission Tomography
SE:
Status Epilepticus
SUV:
Standardized Uptake Value
TLE:
Temporal Lobe Epilepsy
TRITC:
Tetramethyl-Rhodamine Isothiocyanate
TSPO:
18 kDa Translocator Protein
Introduction
Curcumin is a natural polyphenolic yellow pigment extracted from turmeric rhizome
(Curcuma longa L., Zingiberaceae) used for millennia in traditional Indian ayurvedic medicine [1], [2]. Curcumin is a subject of interest in scientific research due to its pleiotropic
therapeutic effects on diseases affecting nearly
every system of the body. Thus, multiple conditions have been claimed to be improved
by curcumin, such as cancer, diabetes, hyperlipidemia, osteoarthritis, myocardial
infarction, different
types of infections, traumatic brain injury, mood disorders, aging, and neurodegenerative
disorders such as Alzheimerʼs, Parkinsonʼs, Huntingtonʼs diseases, and epilepsy [3], [4], [5].
Epilepsy is a neurological disorder that affects more than 50 million people worldwide.
According to the World Health Organization, nearly 80% of epilepsy patients live in
low- and
middle-income countries and it is estimated that 70% of them could live seizure-free
if properly diagnosed and treated (https://www.who.int/news-room/fact-sheets/detail/epilepsy). Among the different types of epilepsy, the temporal lobe epilepsy (TLE) is the
most predominant form of focal epilepsy in
adults [6]. TLE is often accompanied by hippocampal sclerosis [7] and it is highly refractory to the available pharmacological
treatments [8]. Coherently, finding safe, effective, and affordable antiepileptic therapies should
be a main purpose to prevent and/or to counteract the symptoms
therefore reducing the burden associated to this condition [6], [8].
Preclinical studies in different animal models of epilepsy have revealed that curcumin
has anti-seizure and neuroprotective effects, also reducing cognitive impairment [9], [10]. Thus, antiepileptic effects have been reported: (i) delaying the onset of kainic
acid-induced seizures and reducing hippocampal neuronal death
[11]; (ii) preventing iron-induced epileptogenesis [12]; (iii) increasing the threshold current in the electroshock model [13] and (iv) protecting and slowing down the epileptogenic process in both the amygdala
and the pentylenetetrazole kindling models [13], [14].
Pilocarpine, an alkaloid obtained from the leaves of different species from genus
Pilocarpus, is often administered intraperitoneally to trigger status epilepticus (SE), resulting
in
an animal model of epileptogenesis that resembles many, but not all the behavioral,
electrographic, proteomic, and neuropathological features in human TLE [15], [16], [17], being a suitable tool to study the potential antiseizure, antiepileptic and neuroprotective
drugs.
The epileptogenic process in the pilocarpine model, as well as in its variant lithium-pilocarpine
model, is characterized first by the rapid manifestation of the SE, followed by a
latent
silent period without spontaneous seizures. This silent stage is also accompanied
by brain metabolic dysfunction, reflected by a generalized hypometabolism measurable
by
2-deoxy-2-[18F]Fluoro-D-Glucose ([18F]FDG) positron emission tomography (PET) [18]. Brain glucose hypometabolism is concurrent with severe
neurodegeneration and neuronal death, neuroinflammation and intense reactive gliosis,
affecting both astroglia and microglia [19], [20]. Interestingly, in epilepsy patients, glucose hypometabolism during the interictal
period measured by [18F]FDG PET has proven to be very sensitive allowing for the
localization of the epileptogenic focus and its consequences as well as a minimally
invasive procedure [21]. Even though the data regarding the effects of curcumin
on the pilocarpine models are scarce, overall they support its anticonvulsant and
neuroprotective properties [22], [23], [24], [25].
Though curcumin can impact a diverse range of molecular targets and signaling pathways
[5], most of the studies attribute its broad therapeutic benefits primarily
to its antioxidant and anti-inflammatory properties [3], [26]. Because oxidative stress takes part in neuronal damage in epilepsy and
seizures [10], multiple endogenous and exogenous antioxidants have been proposed as add-on therapy.
In fact, it is believed that the antioxidant effect of curcumin
is responsible for protectiion from the pilocarpine-induced SE [24], [27], [28].
While numerous in vitro and in vivo preclinical studies support the potential therapeutic spectrum of curcumin as well
as its safety and tolerability in humans [4], [5], its clinical effectiveness and indication remains to be conclusively confirmed
by randomized, placebo-controlled clinical trials
[10], [29].
To our knowledge, no studies have been conducted in the lithium-pilocarpine SE model
to evaluate the effect of curcumin on the brain metabolic impairment occurring shortly
after the SE,
considered as an early marker of epileptogenesis. Accordingly, herein we used [18F]FDG PET neuroimaging to evaluate brain glucose metabolism during the early phase
of the latent
period of epileptogenesis. Besides, behavioral variables related with SE and different
neurohistochemical assays were performed to evaluate the potential anti-seizure, neuroprotective
and
anti-inflammatory properties of curcumin.
Results
Curcumin did not affect the latency time to reach SE (VEH+PILO: 22.8 ± 2.7 min vs.
CUR+PILO: 19.9 ± 0.6 min, p = 0.275, [Fig. 1 a]). However, curcumin treatment
significantly reduced the number of SE (level 4 – 5 in the Racine scale) that occurred
during the 45 min of observation after the pilocarpine insult (VEH+PILO: 7.0 ± 0.87
vs. CUR+PILO:
3.80 ± 0.70; p = 0.01, [Fig. 1 b]). Death rate reached 50% in VEH+PILO and 58% in CUR+PILO (p = 0.987) reflecting
that curcumin had no effect on the mortality
associated to the SE. Body weight (BW) changes are shown in [Fig. 2 a] and [b]. Compared with their respective controls, VEH+PILO rats
lost a 10% of their BW in the 24 h after the SE (p < 0.01) and this effect remained
until the end of the experiment (4 d) resulting in a total BW loss of 17.4% (d0 to
d4). By contrast,
CUR+PILO rats did not show a statistically significant BW loss 24 h after the SE and
furthermore, they were able to defend their BW, maintaining it throughout the experiment
(CUR+PILO vs.
VEH+PILO, p < 0.05).
 Fig. 1 Curcumin treatment in adult male rats did not delay the onset of SE triggered by
pilocarpine, but it significantly reduced the number of seizures after SE. a Latency
time to the onset of SE. b Number of Racine stage 4 – 5 seizures in VEH+PILO and CUR+PILO. Data are shown as
mean ± SEM (n = 8 – 10 rats/group); *p < 0.05, t-tests
Fig. 1 Curcumin treatment in adult male rats did not delay the onset of SE triggered by
pilocarpine, but it significantly reduced the number of seizures after SE. a Latency
time to the onset of SE. b Number of Racine stage 4 – 5 seizures in VEH+PILO and CUR+PILO. Data are shown as
mean ± SEM (n = 8 – 10 rats/group); *p < 0.05, t-tests
 Fig. 2 SE in adult male rats resulted in a significant BW loss that was prevented by chronic
oral administration of curcumin. a BW data from the day of LiCl administration
(− 1 d) to the day of sacrifice (+ 4 d). b BW changes calculated as percentage of BW on day − 1. Shaded area indicates the 12 h
fasting period before PET acquisitions. Data are
shown as mean ± SEM (n = 3 – 7 rats/group, rats that survived the experimental procedure).
**p < 0.01 VEH+PILO vs. VEH+SAL;
#
p < 0.05 VEH+PILO vs. CUR+PILO;
two-way ANOVA followed by post-hoc Tukey tests
Fig. 2 SE in adult male rats resulted in a significant BW loss that was prevented by chronic
oral administration of curcumin. a BW data from the day of LiCl administration
(− 1 d) to the day of sacrifice (+ 4 d). b BW changes calculated as percentage of BW on day − 1. Shaded area indicates the 12 h
fasting period before PET acquisitions. Data are
shown as mean ± SEM (n = 3 – 7 rats/group, rats that survived the experimental procedure).
**p < 0.01 VEH+PILO vs. VEH+SAL;
#
p < 0.05 VEH+PILO vs. CUR+PILO;
two-way ANOVA followed by post-hoc Tukey tests
An intense hypometabolism in epilepsy-related brain areas was evident 3 days after
the induction of SE ([Fig. 3 a]) as measured by SUV. Compared to VEH+SAL,
VEH+PILO showed a reduction that ranged from 15.4% in hypothalamus to 31.3% in cortex
(p < 0.01; [Fig. 3 b]). Specifically in hippocampus this decrease reached
a 24.8% (p < 0.01; [Fig. 3 b]). Curcumin alone (CUR+SAL) had no significant effect on glucose brain metabolism.
However, curcumin fully prevented SE-induced
hypometabolism ([Fig. 3 b]). Despite the differences in BW, there were not statistical differences among groups
regarding blood glucose concentrations after 12 h
of fasting (measured immediately before PET acquisitions). Thus, the values in mg/dL
were as follows: VEH+SAL: 103.3 ± 3.2; VEH+PILO: 108.0 ± 2.9; CUR+SAL: 105.8 ± 6.3
and CUR+PILO:
104.2 ± 1.9. When brain metabolism was analyzed as [18F]FDG uptake as percentage of the dose injected and corrected by pre-scan whole blood
glucose concentrations
(%IDWBglc), the brain hypometabolism induced by SE was no longer detected (data not shown).
 Fig. 3 SE induced by pilocarpine in adult male rats led to a significant hypometabolism
measured as SUV in key areas known to be involved in epileptogenesis, an effect that
was
ameliorated by curcumin. Regional brain glucose metabolism was evaluated by [18F]FDG PET 3 days after the SE. a Representative CT (upper row), [18F]FDG PET
(mid row) and [18F]FDG PET/CT fused images (bottom row) in coronal, sagittal and trans-axial views
scaled to SUV of the 4 experimental groups. b Regional brain uptake in
the 4 experimental groups is shown as SUV units (mean ± SEM, n = 3 – 7 rats/group,
rats that survived the experimental procedure). *p < 0.05 VEH+PILO vs. VEH+SAL and vs.
CUR+PILO; two-way ANOVA followed by post-hoc Tukey tests
Fig. 3 SE induced by pilocarpine in adult male rats led to a significant hypometabolism
measured as SUV in key areas known to be involved in epileptogenesis, an effect that
was
ameliorated by curcumin. Regional brain glucose metabolism was evaluated by [18F]FDG PET 3 days after the SE. a Representative CT (upper row), [18F]FDG PET
(mid row) and [18F]FDG PET/CT fused images (bottom row) in coronal, sagittal and trans-axial views
scaled to SUV of the 4 experimental groups. b Regional brain uptake in
the 4 experimental groups is shown as SUV units (mean ± SEM, n = 3 – 7 rats/group,
rats that survived the experimental procedure). *p < 0.05 VEH+PILO vs. VEH+SAL and vs.
CUR+PILO; two-way ANOVA followed by post-hoc Tukey tests
Cresyl violet stainings illustrating the qualitative effects of SE reflected an apparent
reduction in hippocampal neurons at the CA1, CA3 and hilus. Curcumin alone (CUR+SAL)
had no effect,
but it seemed to attenuate the effects of SE (CUR+PILO) ([Fig. 4]). These results were in line with and support those obtained from Fluoro-Jade C
fluorescence
labeling. Thus, and as expected, Fluoro-Jade C fluorescence labeling revealed no signs
of neurodegeneration in VEH+SAL or CUR+SAL rats. Instead, SE in VEH+PILO rats triggered
neurodegeneration
in CA1 and hilus (p < 0.01; [Fig. 5 a – b]). Curcumin by itself had no effects but it significantly ameliorated the increase
of Fluoro-Jade C labeling in
CUR+PILO rats ([Fig. 5 a – b]; p < 0.05).
 Fig. 4 Nissl (cresyl violet) staining micrographs from adult male rat brains illustrate,
in a qualitative manner, the damage induced by SE on the anterior hippocampus 3 days
after
the insult. It can be observed that this effect is reduced by curcumin administration.
White arrows indicate the hippocampal areas affected by pilocarpine (CA1, CA3 and
hilus).
Fig. 4 Nissl (cresyl violet) staining micrographs from adult male rat brains illustrate,
in a qualitative manner, the damage induced by SE on the anterior hippocampus 3 days
after
the insult. It can be observed that this effect is reduced by curcumin administration.
White arrows indicate the hippocampal areas affected by pilocarpine (CA1, CA3 and
hilus).
 Fig. 5 Hippocampal neurodegeneration induced by SE is ameliorated by chronic curcumin treatment
in adult male rats. a Representative images at the level of the CA1 (top
row), CA3 (mid row) and dentate gyrus/hilus (bottom row) of the 4 experimental groups.
The images show degenerating neurons in VEH+PILO rats 3 days after the SE over an
increased FITC
fluorescence background signal, and the protective effect of curcumin (CUR+PILO).
b Bar plot corresponding to quantitative data from Fluoro-Jade C fluorecence intensity
values as
marker of neurodegeration. Data are expressed as percentage of the signal obtained
in the VEH+SAL group and shown as mean ± SEM (n = 3 – 7 rats/group; rats that survived
the experimental
procedure). **p < 0.01 VEH+PILO vs. VEH+SAL and vs. CUR+PILO; two-way ANOVA followed
by post-hoc Tukey tests
Fig. 5 Hippocampal neurodegeneration induced by SE is ameliorated by chronic curcumin treatment
in adult male rats. a Representative images at the level of the CA1 (top
row), CA3 (mid row) and dentate gyrus/hilus (bottom row) of the 4 experimental groups.
The images show degenerating neurons in VEH+PILO rats 3 days after the SE over an
increased FITC
fluorescence background signal, and the protective effect of curcumin (CUR+PILO).
b Bar plot corresponding to quantitative data from Fluoro-Jade C fluorecence intensity
values as
marker of neurodegeration. Data are expressed as percentage of the signal obtained
in the VEH+SAL group and shown as mean ± SEM (n = 3 – 7 rats/group; rats that survived
the experimental
procedure). **p < 0.01 VEH+PILO vs. VEH+SAL and vs. CUR+PILO; two-way ANOVA followed
by post-hoc Tukey tests
In the VEH+PILO group, the SE resulted in an approximately 254% increase in fluorescence
GFAP signal in the hippocampal hilus (vs. VEH+SAL group, p = 0.003; [Fig. 6 a – b]), pointing towards SE-induced astroglia activation. Compared to VEH-PILO rats, curcumin
reduced the astrocytic activation in response to SE. In this way, the quantitative
data revealed that the fluorescence signal found in CUR+PILO in response to SE was
approximately a 31% lesser than in VEH+PILO group (p = 0.039). Besides, and as it
can be observed in [Fig. 6 c], the astroglia activation was accompanied by a qualitative apparent thickening of
astrocyte bodies and processes.
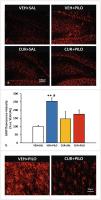 Fig. 6 Curcumin reduced hippocampal astrogliosis in response of the SE triggered by pilocarpine
in adult male rats. a Images showing representative GFAP immunofluorescence
micrographs at the hilus/dentage girus area of the 4 experimental groups. b Bar plot corresponding to quantitative data from GFAP immunofluorecence intensity
as marker of
astrogliosis on the hilus. Data are expressed as percentage of the signal obtained
in the VEH+SAL group and shown as mean ± SEM (n = 3 – 7 rats/group; rats that survived
the experimental
procedure). **p < 0.01 VEH+PILO vs. VEH+SAL and
#
p < 0.05 VEH+PILO vs. CUR+PILO; two-way ANOVA followed by post-hoc Tukey tests. c Magnified micrographs
illustrate the apparent morphological changes (hypertrophy of astrocyte bodies and
processes) induced by SE and the effect of curcumin.
Fig. 6 Curcumin reduced hippocampal astrogliosis in response of the SE triggered by pilocarpine
in adult male rats. a Images showing representative GFAP immunofluorescence
micrographs at the hilus/dentage girus area of the 4 experimental groups. b Bar plot corresponding to quantitative data from GFAP immunofluorecence intensity
as marker of
astrogliosis on the hilus. Data are expressed as percentage of the signal obtained
in the VEH+SAL group and shown as mean ± SEM (n = 3 – 7 rats/group; rats that survived
the experimental
procedure). **p < 0.01 VEH+PILO vs. VEH+SAL and
#
p < 0.05 VEH+PILO vs. CUR+PILO; two-way ANOVA followed by post-hoc Tukey tests. c Magnified micrographs
illustrate the apparent morphological changes (hypertrophy of astrocyte bodies and
processes) induced by SE and the effect of curcumin.
We also performed [3H]PK11 195 autoradiography as a marker of neuroinflammation at the level of the anterior
and posterior hippocampus ([Fig. 7 a]).
Curcumin by itself had no effects on [3H]PK11 195 binding. Regarding the effects of pilocarpine-triggered SE and comparing
it to VEH+SAL, VEH+PILO rats showed an approximately 100%
increase in the optical density (O. D.) in all the regions studied (p < 0.01; [Fig. 7 b]). This difference in signal was also found between VEH+PILO and
CUR+PILO groups (p < 0.01; [Fig. 7 a – b]). These effects were also significant when measured and analyzed in the hippocampal
CA1, CA3 and hilus subregions
([Fig. 7 c]).
 Fig. 7 Chronic curcumin treatment reduced the neuroinflammation induced by SE in adult male
rats as measured by the [3H]PK11 195 binding in major brain regions involved
in epileptogenesis. a Representative [3H]PK11 195 autoradiograms corresponding to the 4 experimental groups obtained from
both anterior and posterior hippocampus.
b [3H]PK11 195 binding expressed in O. D. in various brain regions involved epileptogenesis.
c [3H]PK11 195 binding expressed in O. D. in the hipocampal
regions CA1, CA3 and hilus. Data is shown as mean ± SEM (n = 3 – 7 rats/group; rats
that survived the experimental procedure). *p < 0.05 and **p < 0.01 VEH+PILO vs. VEH+SAL
and vs.
CUR+PILO; two-way ANOVA followed by post-hoc Tukey test
Fig. 7 Chronic curcumin treatment reduced the neuroinflammation induced by SE in adult male
rats as measured by the [3H]PK11 195 binding in major brain regions involved
in epileptogenesis. a Representative [3H]PK11 195 autoradiograms corresponding to the 4 experimental groups obtained from
both anterior and posterior hippocampus.
b [3H]PK11 195 binding expressed in O. D. in various brain regions involved epileptogenesis.
c [3H]PK11 195 binding expressed in O. D. in the hipocampal
regions CA1, CA3 and hilus. Data is shown as mean ± SEM (n = 3 – 7 rats/group; rats
that survived the experimental procedure). *p < 0.05 and **p < 0.01 VEH+PILO vs. VEH+SAL
and vs.
CUR+PILO; two-way ANOVA followed by post-hoc Tukey test
Discussion
In the current work, we have explored the effects of chronic oral administration of
curcumin on brain glucose metabolic dysfunction, hippocampal neurodegeneration, and
neuroinflammation,
typical features of the brain damage associated to the SE in the rat lithium-pilocarpine
model [19]. We have also studied the effects of curcumin on latency to SE,
number of seizures, mortality, and BW change. Overall, our results show that curcumin
did not affect either latency time to SE or mortality rate. However, curcumin treatment
significantly
reduced the number of stage 4 – 5 seizures and ameliorated signs of brain damage associated
with the SE also having a marked protective effect on BW in the rats that survived
the insult.
Curcumin is a polyphenol extracted from the rhizomes of Curcuma longa (family Zingiberaceae). Commonly known as turmeric and referred to as the “golden
spice” and “spice of life”,
curcumin has been traditionally employed as a dietary component as an herb, a spice,
as a cosmetic product, and as a natural medicinal agent in Asia, particularly in Ayurveda
medicine [1], [2]. In fact, numerous in vitro and in vivo preclinical and clinical studies on curcumin have put forth pleiotropic
beneficial effects dealing with its anti-cancer, anti-diabetic, antimicrobial, antioxidant,
and anti-inflammatory properties [30]. Furthermore, curcumin has been
reported to have neuroprotective and cognitive-improving properties that may delay
or prevent many of the deleterious processes occurring in most neurodegenerative and
neurological diseases,
including epilepsy [31].
Despite these alleged beneficial effects, it is important to mention that recent studies
have called into question the real in vivo effectiveness of curcumin. One of the main
limitations is the poor physical-chemical and pharmacokinetic properties of the curcumin
molecule characterized by low aqueous solubility, gut absorption, and limited entry
to the CNS through
the blood brain barrier, as well as rapid metabolism and systemic elimination [9], [29], [31], [32].
In keeping with the controversy, curcumin, like many other natural compounds, has
been labeled as both PAINS (pan assay interference compounds) and IMPS (invalid metabolic
panaceas) compounds
[29]. Thus, curcumin might interfere with many in vitro and ex vivo biochemical assays used to evaluate multiple biological activities, leading to
erroneous claims for a non-existent therapeutic effect. Yet, despite these caveats,
the overwhelming amount of wide-range experimental evidence regarding the beneficial
effects of curcumin,
including its potential therapeutic role in epilepsy, cannot be set aside.
In the present study, and to tackle the poor oral bioavailability of curcumin, we
used 10% Cremophor EL (10 mL/kg) as vehicle [25], [33]. Cremophor EL is a non-ionic solvent for hydrophobic compounds that has been shown
to improve solubility [34] and oral bioavailability of curcumin [35]. Even though Cremophor EL is a non-inert relatively nontoxic solvent, it is important
to mention that several reports suggest that it can induce serious
complications such as anaphylactoid-hypersensitivity and cellular toxicity, especially
when administered intravenously [36]. Nevertheless, observational follow-up
of our rats, including BW changes, throughout the whole duration of our study, indicated
neither digestive nor other signs of toxicity.
As previously mentioned, the SE induced by lithium-pilocarpine is an animal model
of epileptogenesis that resembles many, but not all, pathological features of human
TLE [15], [16], [17]. In so far, to date few studies have tackled the therapeutic efficacy of curcumin
(administrated either as a single dose or repeatedly) on the pilocarpine model of
SE in rats [24], [25], [27], [28], [37]. Epileptogenesis is associated, among others, with neurochemical imbalances, neurodegeneration,
neuroinflammation, and reactive gliosis, as well as synapsis modification and reorganization
of specific brain areas [19], [38]. Many
of these alterations are present in the rodent pilocarpine and lithium-pilocarpine
SE models [6], [16], [17]. In these models, pilocarpine administration results in a SE that is followed by
a silent latency period during which generalized glucose hypometabolism occurs concomitantly
with
severe neurodegeneration and neuronal death, neuroinflammation, and intense reactive
gliosis, affecting both astroglia and microglia [18], [19], [20], and ultimately leading to a chronic epileptic state characterized by spontaneous
recurrent seizures. Therefore, it is in this period that
[18F]FDG PET acquisitions and neurohistochemical assessments were carried out.
In our study, curcumin treatment neither delayed the latency time to the SE nor reduced
the mortality rate consequence of the severity of SE induced by pilocarpine. The mortality
rate in our
current study, being around 50%, is within the range for this model. In fact, the
death rate described for the rat lithium-pilocarpine model can be as high as 95%.
Furthermore, high
intra-strain, inter-strains and sub-strains variability have been linked to different
pilocarpine sensitivity. Even more, the commercial providers and the time of purchase
of animals seem to
be factors contributing to the variability on mortality [39]. However, curcumin significantly reduced the number of stage 4 – 5 seizures that
occurred during the
45 min after the beginning of the SE. Unfortunately, we did not measure the duration
of the seizures. Nevertheless, our results might point towards an anticonvulsant effect
of curcumin that
would be in line with previous reports [28], [37]. Likewise, anticonvulsant effects of oral curcumin have been also reported in other
animal models of epilepsy and seizures such as the iron-induced experimental model
of epileptogenesis in rats [12] and in the pentylenetetrazole-kindled rat model
of epilepsy [40], [41]. Therefore, it is likely that the effects of curcumin reducing the number of seizures
might be one the factors
contributing to the control of further spontaneous seizures and to the neuroprotective
effects observed in the rats that survived the SE [41], [42]. Nonetheless, lack of long-lasting anti-epileptogenic, neuroprotective and anti-inflammatory
effects of intracerebroventricular administration of curcumin have been
also reported in a kindling rat model [43].
BW change is widely accepted as a marker of the overall animal well-being. In rats
that did not undergo SE, oral curcumin administration did not affect BW change throughout
the 17 days of
experimental procedure. By contrast, curcumin significantly reduced the effects of
SE inducing BW loss ([Fig. 2]). Thus, curcumin treatment allowed rats to defend
their BW in the face of the SE. It is likely that the effect of curcumin enhancing
the ability of the rats to defend their BW is both a reflection of the reciprocal
central and peripheral
protective effects of curcumin that, could ultimately contribute to set in motion
a more adaptive response to the metabolic demands imposed by the SE. In this line
of reasoning, other studies
in rats have shown that curcumin treatment protected from BW loss promoting resilience
to chronic social defeat stress [44], and reduced BW loss in response to
2,3,7,8-tetrachlorodibenzo-p-dioxin administration [45]. Interestingly, curcumin also has shown beneficial effects on BW reduction and energy
metabolism in rodent
models of obesity, nowadays accepted as a pro-inflammatory disease [46].
The interictal temporal lobe glucose hypometabolism is one of the early biomarkers
identified by [18F]FDG PET neuroimaging in TLE patients, providing even better results than
magnetic resonance imaging (MRI) [47], [48]. The hypometabolism has been attributed, among others, to neuronal death, altered
neuronal
excitability, and/or reduced brain blood flow in the epileptic focus. Importantly,
brain glucose hypometabolism has been repeatedly reproduced in many animal models,
including the pilocarpine
model [19], [49], [50], [51]. Our current study corroborates that the SE
induced by pilocarpine results in glucose hypometabolism (measured by SUV) in epilepsy-related
brain areas ([Fig. 3]).
More remarkably, glucose hypometabolism was fully prevented by curcumin treatment
([Fig. 3]). As far as we know, this is the first time that functional
neuroimaging PET has been implemented to assess the effect of curcumin on glucose
hypometabolism induced by SE.
Considering that brain glucose hypometabolism seems to be an early biomarker of epileptogenesis
in different animal models of epilepsy [52], our results might
point towards an antiepileptogenic effect of curcumin in this model. Nevertheless,
because we have not evaluated the long-term effects, we cannot state that the effect
of curcumin preventing
SE-triggered brain glucose hypometabolism is necessarily associated with the alleged
antiepileptic effect.
Nonetheless, we want to notice that the characteristic brain glucose hypometabolism
in response to SE was not observed when quantified as percentages of the injected
dose and corrected by the
pre-scan whole blood glucose values (%IDWBglc). It is known that SUV and %ID need to be corrected for glucose, especially when
the fasting period is shorter than 12 h [53]. Moreover, not considering the rapid BW loss in response to SE (see [Fig. 2]) might result in underestimation in hypometabolism
quantification [53], [54].
In agreement with previous reported studies, pilocarpine-triggered SE resulted in
hippocampal neuronal death and neurodegeneration [19]. The results obtained
regarding hippocampal integrity based on the visual observation of Nissl staining,
and neurodegeneration based on the quantitation of Fluoro-Jade-C fluorescence [55], [56], support a neuroprotective effect of curcumin treatment. Thus, curcumin contributed
to preserve neuronal integrity in the CA1 and hilus,
hippocampal areas where the apparent neuronal loss ([Fig. 4]) and marked neurodegeneration ([Fig. 5]) occurred in response to
pilocarpine. Those results support the neuroprotective effects that have been attributed
to curcumin, mainly based on its antioxidant and anti-inflammatory properties [14], [57]. Similarly, a previous study in the lithium-pilocarpine model has reported that
curcumin protected hippocampal neurons through induction of
autophagy and inhibition of necroptosis [25].
Neuroinflammation is an adaptive physiological response to brain cell damage or loss
and, gliosis affecting both astrocytes and microglia have been reported in epileptic
disorders. We and
others have consistently reported that neuroinflammation is also a feature of the
pilocarpine rodent model [19], [20] as well as of
other animal models [49], [58]. Accordingly, our current results show that SE triggered reactive astrogliosis and
microglia-mediated
neuroinflammation. Furthermore, our results reveal that curcumin significantly ameliorated
both hippocampal astrogliosis (evaluated by GFAP immunofluorescence; [Fig. 6]) and brain microglia-mediated neuroinflammation (evaluated by [3H]PK11195 autoradiography). PK11195 is a ligand of the mitochondrial 18 kDa translocator
protein
(TSPO), which is mainly, but not exclusively, present in microglia. Thus, TSPO is
expressed in vascular endothelial cells and astrocytes. Brain TSPO expression is low
under non-pathological
conditions, but it is upregulated in response to neuroinflammation, being considered
as a marker of neuroinflammation in many neurological diseases [59]. In our
study, [3H]PK11195 autoradiography did not include non-specific binding, as such. It has been
reported that curcumin can interact directly with low micromolar affinity with TSPO
[60]. This might point towards a potential direct anti-inflammatory mechanism of action
for curcumin. Nevertheless, because the binding was performed in brains
collected 2 days after the last curcumin administration, it is most likely that the
48 h washout period, added to the poor pharmacokinetic properties of curcumin, may
have significantly
reduced these eventual binding interferences in all curcumin-treated rats. Herein,
we show that SE results in an increase in [3H]PK11195 binding signal ([Fig. 7]). This increase is in line with previous studies reporting increased [18F]GE180 (a TSPO PET tracer) signal when neuronal damage occurs as consequence of seizures
and
epileptogenesis [19], [49]. Altogether, these results point towards an anti-inflammatory and neuroprotective
effect of oral
curcumin.
Numerous and redundant are the mechanisms attributed to curcumin neuroprotective properties,
including its ability to reduce oxidative stress and to regulate anti-inflammatory
and
pro-inflammatory pathways [61]. Despite its low bioavailability and the questionable blood brain barrier-crossing
previously mentioned [29], various mechanisms might partially explain the central effects of curcumin in brain.
Among them, we can mention the implication of the gut-brain axis, involving the metabolic
role
of microbiota [31] as well as upregulation of epithelial enzymes with antioxidant and anti-inflammatory
effects [62]. Besides, it is
interesting to mention that curcumin improves ghrelin expression [63], [64]. Furthermore, in the pilocarpine rat model of SE, ghrelin
and ghrelin analogs have shown to be neuroprotective without anticonvulsant effects
[65].
To summarize, chronic (17 d) oral curcumin treatment neither delayed the SE nor reduced
the mortality rate associated with our experimental model. However, curcumin seemed
to have a certain
anti-seizure effect that, in the survival rats, might have contributed to the amelioration
of the damage induced by SE. Actually, BW loss, brain glucose hypometabolism, neurodegeneration,
and
neuroinflammation were reduced by curcumin treatment. Furthermore, and as far as we
know, this is the first time that functional neuroimaging PET has been implemented
to assess the effect of
curcumin on brain glucose hypometabolism induced by SE. Although the exact direct
and/or indirect mechanisms need further enquiry, and despite the controversy regarding
the actual
bioavailability and effectiveness of oral curcumin, our overall results would support
the claimed neuroprotective and anti-inflammatory properties of this phytochemical.
Altogether, taking
into consideration that curcumin in the absence of injury had no effect, it is likely
that curcumin has adaptogen-like properties, enabling the animals to withstand and
to adaptatively respond
to the demands imposed by insults of various nature.
Materials and Methods
Animals and drug treatment protocol
Adult male Sprague-Dawley rats (Charles River Laboratories) weighing 326.9 ± 3.5 g
at the beginning of the experiment were used. Rats were housed in standard rat cages
(2 rats/cage), on a
ventilated rack (Tecniplast) under controlled temperature (22 ± 2 °C) and a 12 h light/dark
cycle (8 : 00 AM-8:00 PM). Throughout the study, rats had free access to standard
rodent chow and
tap water, were weighed daily at morning and BW registered as a marker of overall
welfare. Food was removed for the 12 h before the [18F]FDG PET acquisitions to reduce competition
between plasma glucose and the radiotracer for the glucose transporters. The study
was approved on the 10th of August of 2015 by both the Animal Research Ethical Committee
of the Universidad
Complutense de Madrid and by the Autonomous Community of Madrid (PROEX 238/15), being
carried out in accordance with regulations of the European Union (2010/63/UE) and
Spain (RD53/2013)
regarding animal welfare.
Four experimental groups were used in the present study: (1) control naïve group:
rats received Cremophor EL daily as vehicle instead of curcumin (14 + 3 days) and
saline instead of
lithium-pilocarpine, therefore they never underwent SE (VEH+SAL); (2) rats that received
Cremophor EL daily as vehicle instead of curcumin (14 + 3 days) and underwent
lithium-pilocarpine-induced SE group (VEH+PILO); (3) rats that received curcumin (14 + 3
days) and were not exposed to SE (CUR+SAL) and finally, (4) rats that chronically
received curcumin
and underwent the lithium-pilocarpine insult (CUR+PILO). Besides all rats were exposed
to the same procedures required for PET studies.
SE induction
The lithium-pilocarpine model of SE followed in the current study has been described
in previous studies [19]. Briefly, lithium chloride (127 mg/kg i. p.,
Sigma–Aldrich) was administered 18 – 20 h before SE induction. The next day, and following
the daily protocol, either curcumin or vehicle were administered at morning. Approximately
2 h
later, methyl-scopolamine (2 mg/kg, i. p.) was administered to reduce the pilocarpine-induced
muscarinic peripheral effects. Thirty min later, pilocarpine was injected (25 mg/kg,
i. p.;
Sigma–Aldrich). The onset of SE was considered when the animal reached the stage 4
according to the Racine scale [66] and showed continuous seizure activity. The
seizure activity was ended by injecting pentobarbital (25 mg/kg, i. p.) 45 min after
the SE onset. The rats that did not undergo SE (naïve- and curcumin-control groups)
were administered
with the same drug regime (including lithium chloride, methyl-scopolamine, and pentobarbital)
but saline solution was administered instead of pilocarpine.
Curcumin treatment
Because the relevance of preclinical studies mainly rely on their prospective eventual
translation into the clinical setting, we administered curcumin by oral route, by
intragastric gavage.
To improve bioavailability, curcumin was suspended in 10% Cremophor EL (Sigma-Aldrich)
[25], [33]. Curcumin was administered once
daily in the morning (300 mg/kg/10 mL, p. o.) for 14 days before the SE, the day of
the SE induction and for the 2 following days. The dosing regime was chosen based
on a previously reported
study administering either 200 or 300 mg/kg of curcumin for 14 days, by intragastric
gavage, to adult Sprague-Dawley male rats in the lithium-pilocarpine model of SE [25]. To prevent any eventual degradation of curcumin, the suspension was freshly prepared
every morning and the container wrapped with aluminum foil to protect it from light.
According to the 3Rs principles and the ARRIVE guidelines (https://arriveguidelines.org), the number of animals was selected considering the
mortality associated with the severity of the model. Besides, considering that: (i)
studies do not support for relevant effects of curcumin under baseline or control
conditions; (ii) main
effects of curcumin have been reported when animals were exposed to psychological,
physical, or chemical insults and, (iii) our current main objective was to study the
potential effects of
curcumin in the face of the insult triggered by SE, the number of animals in the control
groups was reduced. The initial experimental sample size was then as follows: (1)
VEH+SAL, n = 3; (2)
VEH+PILO, n = 14; (3) CUR+SAL, n = 4; and (4) CUR+PILO, n = 14. After the mortality
caused by the pilocarpine treatment the final sample size was: (1) VEH+SAL; n = 3;
(2) VEH+PILO; n = 7;
(3) CUR+SAL; n = 4; and (4) CUR+PILO; n = 6.
[18F]FDG PET neuroimaging
PET scans were carried out 3 days after the SE. To this aim, a hybrid PET/CT (computed
tomography) scanner (Albira scanner, Bruker NMI) was used. The protocols have been
previously reported
[19], [49]. Briefly, the rats were fasted (12 h) before scanning. [18F]FDG was injected into the tail vein (approximately
13 MBq − 350 µCi- in 0.2 mL of 0.9%; Curium Pharma) and 30 min later, PET and CT acquisitions
were consecutively carried out under isoflurane anesthesia. After reconstruction of
the
tomographic images, the metabolic activity was quantified using PMOD 3.6 software
(PMOD Technologies Ltd.). As index of metabolic activity, the standardized uptake
value (SUV) for each
region was then calculated, taking into account the rat weight, the dose injected,
and the [18F]FDG uptake decay-corrected to the scan start time. Nevertheless, and because the
effects of pilocarpine and/or curcumin on brain glucose metabolism might be indirectly
reflecting changes in BW and/or glucose metabolism, we also report the regional brain
[18F]FDG uptake data as %IDWBglc
[53], [54].
Neurohistochemical assessments
Rats were sacrificed by decapitation the day after the PET acquisition procedure.
Brains were dissected, cut longitudinally in two halves, quickly frozen on dry ice,
and stored at − 80 °C.
Brain slices (30 µm-thickness) from the left hemibrain were obtained using a cryostat
(Leica CM1850, Leica Biosystems). The brain sections containing the hippocampus (6
slices/glass slide)
were thaw-mounted onto Superfrost Plus slides (Thermo Scientific), dried on a hot
plate (36 °C) and stored into slide boxes at − 80 °C until the day of the assays.
The histochemical assays
were as follows:
-
Neuronal viability and disruption of hippocampal integrity was evaluated by Nissl staining as previously described [19], [49]. Briefly, the slices were fixed in 4% formaldehyde in phosphate buffer pH 7.4 (10 min),
washed, and incubated for 30 min in 0.5% cresyl violet acetate solution. Afterwards,
the sections were washed and dehydrated in graded ethanol series (70%, 95% and 100%).
Finally, the slices were cleared in xylene and cover-slipped with DPX mounting medium
(Fluka). The
histological images of the hippocampus were captured with a digital camera (Leica
DFC425, Leica) coupled to a microscope (Leica DM 2000 LED, Leica). The histological
images are presented
as a visual aid reflecting the qualitative changes induced by SE and the potential
effect of curcumin on the hippocampus.
-
Hippocampal neurodegeneration was evaluated by Fluoro-Jade C staining, as previously reported [19], [49], [56]. Briefly, after fixing in 4% formaldehyde for 10 min, the samples were rinsed in
basic alcohol, 100% ethanol, distilled water, 0.06% potassium permanganate,
0.1% acetic acid solution containing 0.0001% Fluoro-Jade C (Millipore), distilled
water, and xylene. Next, the slides were cover-slipped with DPX (Fluka). The fluorescence
images were
captured with a digital camera (Leica DFC3000G) coupled to a microscope (Leica DM
2000 LED) by using the FITC filter. At the hippocampal CA1, CA3 and hilus, the fluorescence
signal was
measured using ImageJ 1.46 r software. The average value for each rat was calculated.
The results are expressed as percentage vs. the control group (VEH+SAL).
-
Reactive astrogliosis was evaluated by glial fibrillary acidic protein (GFAP) one-step immunofluorescence
as previously reported [19], [49]. Briefly, after fixing and washing, the slices were blocked and permeabilized with
3% BSA, 0.1% triton X-100 in TBS for 60 min. The slides were then incubated
overnight with anti-GFAP-Cy3 antibody (1 : 500, Sigma Aldrich) in 1% BSA in TBS at
4 °C. Afterwards, the slides were washed in 0.1% Tween 20 dissolved in Tween for 3
times (5 min each)
and cover-slipped with Mowiol. The images were captured and examined using the same
optical systems used for Fluoro-Jade C with the TRITC filter. For each brain section
(4 sections/rat)
containing the CA1, CA3 and hilus areas within the anterior (dorsal) hippocampus were
selected and the fluorescence intensity was measured (ImageJ 1.46 r software). The
average value for
each rat was calculated. The results are expressed as percentage vs. the control group
(VEH+SAL).
-
Neuroinflammation was studied by [3H]PK11 195 autoradiography with minor modifications [67]. [3H]PK11 195 is a specific ligand
of the 8 kDa translocator protein (TSPO), previously known as the peripheral benzodiazepine
receptor. TSPO is predominantly, but not exclusively, expressed in microglia, and
increases in
conditions of neuroinflammation [59]. Slides were dried at 37 °C (10 min) and preincubated with 50 mM Tris-HCl pH 7.4
at RT (15 min). The samples were then
incubated with 1 nM [3H]PK11 195 (Perkin Elmer) in preincubation buffer (60 min). Afterwards, the samples
were washed in ice cold preincubation buffer (2 × 5 min) and dipped
in ice-cold distilled water. Once dry, the slides were exposed to Kodak BioMax MR
autoradiography film (Carestream) inside an exposure cassette for 2 months. The developed
film was
placed onto a light box (Kaiser Prolite 5000, Kaiser Fototechnik) and the images from
each section were captured with a camera (Leica DFC425) coupled to a stereomicroscope
(Leica MZ6).
Within each brain section, the O. D. was measured in the selected regions and in the
background. The O. D. values obtained after subtracting the background were expressed
as percentage
of the VEH+SAL group and used as index of neuroinflammation.
Statistical analyses
Analyses were performed with SigmaPlot 11.0 software (Systat Software Inc.). Behavioral
markers of SE onset (latency), number of seizures, and mortality rate were only analyzed
in the
lithium-pilocarpine-treated rats (VEH+PILO vs. CUR+PILO) by unpaired Student t-test
and z-test for rates and proportions, respectively. BW, PET, and histochemical data
were analyzed by
two-way analysis of variance (ANOVA) with curcumin and pilocarpine treatments as the
two main factors. When interaction between factors was significant, further post hoc
Tukey tests were
performed. In all cases, statistical significance was considered when p < 0.05. Data are shown as mean ± SEM.
Contributorsʼ Statement
Data collection: L. García-García, M. Delgado, R. Fernández de la Rosa, K. Slowing
Design of the study: L. García-García, F. Gomez, M. Delgado, K. Slowing Statistical
analysis: L.
García-García, M. Delgado, N. Hernández-Martín Analysis and interpretation of the
data: F. Gomez, L. García-García Drafting of the manuscript: F. Gomez, L. García-García
Critical revision:
M. A. Pozo





