Keywords
amelogenin - FABP-3 - fetal mice - enamel density - nanoparticle - saltwater fish
powder
Introduction
Enamel, the outermost protective layer of teeth, serves as the first line of defense
against mechanical and microbial insults. Its formation, known as amelogenesis, occurs
predominantly during fetal development and is critical for determining the long-term
resilience of dental tissue.[1] Enamel is particularly susceptible to caries and structural defects if its quality
and density are compromised, emphasizing the importance of understanding factors that
influence its development.[2] Amelogenesis is generally divided into five stages: the pre-ameloblast, pre-secretory,
secretory, transition, and maturation phases. These stages are characterized by morphological
and functional changes in ameloblasts, a monolayer of specialized epithelial cells
responsible for enamel matrix secretion and subsequent mineralization.[3] During the maturation phase, significant mineral deposition occurs, leading to the
removal of the organic matrix components and increased enamel hardness and density.[4]
[5]
Amelogenin is a key protein involved in dental enamel formation[6] and plays a crucial role in regulating hydroxyapatite crystal growth and organization,
thus directly affecting the enamel mineralization density and mechanical structure.[7]
[8] Fatty acid-binding proteins (FABPs), particularly FABP-3, are lipid transporters
implicated in cellular lipid metabolism, which has been suggested to influence ameloblast
function and enamel mineralization through modulation of lipid availability and signaling
pathways.[9] Although the direct involvement of FABP-3 in enamel-forming cells has not yet been
fully established, its known role in lipid handling highlights a potential contribution
to enamel biomineralization, warranting further investigation into its specific function
in ameloblasts.
Marine biota, especially saltwater fish, are rich sources of essential nutrients such
as protein, minerals, and bioactive compounds including omega-3 fatty acids, calcium,
vitamin A, D, and B12, sodium, fluoride, and selenium.[10] These nutrients are known to influence key pathways involved in enamel development.
For instance, omega-3 fatty acids like eicosapentaenoic acid (EPA) and docosahexaenoic
acid (DHA) support lipid metabolism and have been shown to promote FABP-3 expression,
facilitating lipid metabolism and mineralization during amelogenesis.[11] Additionally, amino acids like histidine and glycine are fundamental constituents
of amelogenin, influencing its synthesis and functionality. Vitamins and minerals
such as vitamin D, calcium, and phosphorus are vital for hydroxyapatite crystal formation.[12]
[13]
[14] However, the bioavailability of nutrients from natural dietary sources is often
limited due to factors such as molecular and absorption insufficiencies, such as low
solubility, limited intestinal absorption, and restricted placental transfer.
Recent advances in nanotechnology offer promising opportunities to enhance nutrient
delivery.[15] Nanoparticle formulations, ranging from 50 to 300 nm in size, increase bioavailability
by facilitating better absorption and cellular uptake of minerals and bioactive compounds
such as calcium, phosphate, and omega-3 fatty acids.[16] Applying nanoparticle technology to nutrient delivery could potentiate the effects
of marine-derived nutrients on amelogenesis, yet scientific evidence on their specific
impact on fetal enamel formation remains limited.
Therefore, this study aims to investigate the effects of nanoparticle saltwater fish
powder supplementation on the expression of key enamel proteins (amelogenin) and FABP-3,
in fetal mice. FABP-3 was assessed in the placenta as an indicator of the ability
of nanoparticles to cross the placental barrier. Furthermore, this research evaluates
the influence of this intervention on enamel density using micro-computed tomography
(micro-CT/μCT), contributing to the understanding of nutritional strategies for optimal
fetal enamel development.
Materials and Methods
Study Design
This randomized experimental study was conducted using a single-blind design in which
the animals were randomly allocated into control and treatment groups. The researchers
administering the intervention were aware of group assignments, while the outcome
evaluators remained blinded to reduce the risk of bias. The study was approved by
the Research Ethics Committee of the Faculty of Dentistry, Universitas Islam Sultan
Agung, Semarang (approval no. 603/B.1-KEPK/SA-FKG/IX/2024).
Preparation of Nanoparticles from Saltwater Fish Powder
Nanoparticle saltwater fish powder was prepared from sardines (Sardinella fimbriata), splendid pony fish (Leiognathus splendens), and tuna (Euthynnus affinis). The fish were dried and ground into powder using an Ultra Turrax (IKA, Germany).
Semi-quantitative XRF (SQX) analysis of the saltwater fish powder sample revealed
that the dominant elements were calcium (Ca) at 8.50 mass% and phosphorus (P) at 3.68
mass%, with a calculated Ca/P ratio of approximately 2.31. Gas chromatography–flame
ionization detector (GC-FID) analysis, using method code 18-6-1/MU/SMM-SIG, revealed
that the saltwater fish powder nanoparticles contained 74.2 mg/100 mg of omega-3,
647.6 mg/100 mg of omega-6, and 16,675.5 mg/100 g of omega-9 fatty acids.
The powder was mixed with nanochitosan dissolved in 2% acetic acid and sodium tripolyphosphate,
then homogenized using a magnetic stirrer (IKA, Germany). Particle size analyzer analysis
using the dynamic light scattering method revealed a particle size distribution ranging
from approximately 61 to >7,000 nm, with several main distribution peaks. The sample
exhibited a nanoparticle fraction at approximately 91 nm, with a Z-average of 166.7 nm
and an average of 273.3 nm, along with a particle size distribution extending into
the micrometer range.
Experimental Animals
Healthy mice (Mus musculus) displaying normal behavior without lethargy or wounds were selected. A total of 16
female mice and 8 males (weighing 20–30 g, aged 2–3 months) were used. The mice were
housed under controlled conditions (temperature: 23–27°C, light-dark cycle: 12 hours)
with ad libitum access to food and a standard laboratory rodent diet (commercial pellet
feed). Female mice's estrous cycles were monitored via vaginal swabs, and those in
estrus were mated with males (ratio: four females to two males). A vaginal plug confirmed
day 0 of gestation.
In addition to ad libitum access to a standard rodent pellet diet and water, the treatment
group received oral administration of saltwater fish powder nanoparticles (2.14 mg/mL)
dissolved in distilled water three times daily, while the control group received the
same volume of distilled water (vehicle control) for 17 days.
Sample Collection and Analysis
On gestational day 18, the mice were sedated with 10 to 20 mL of chloroform. Respiratory
and heart rates were monitored before cervical dislocation. Placenta and fetal jaws
were collected, fixed in 10% formalin, and processed for histological and immunohistochemical
(IHC) analysis. The human protein atlas database was used as a reference for antibody
selection and staining protocols to ensure specificity for murine tissues.
Amelogenin and FABP-3 Expression Evaluation
IHC analysis was performed to evaluate the expression of amelogenin, a key marker
of enamel matrix formation during odontogenesis in the developing tooth buds of fetal
mice, and FABP-3 marker of transport and accumulation of fatty acids in placental
tissues. Tissue sections were stained to visualize amelogenin and FABP-3 distribution
in the tooth bud and placental tissues, respectively, and examined under a light microscope.
A mouse monoclonal antibody against amelogenin (F-11):sc-365284 (Santa Cruz Biotech
Inc., Delaware, California, United States) was used as the primary antibody. A rabbit
polyclonal FABP3 bs-11283R-HRP (Bioss ANTIBODIES, Boston, Massachusetts, United States)
was used as the conjugated primary antibody. Amelogenin expression was measured 10
times per sample using a light microscope (Leica DM 750, Germany) at magnifications
of 100× and 400×. Meanwhile, the FABP-3 analysis was conducted on 10 fields per sample
of placental tissue.
Enamel Density Evaluation
Enamel density was measured using µCT (SkyScan1173; Bruker, Kontich, Belgium). The
µCT analysis was conducted by examining the average enamel density through sagittal,
axial, and anteroposterior sections. Eight fetal mouse samples were used in both the
control and treatment groups. The system used cone beam X-ray transmission with a
flat-panel CCD camera (50 µm pixel size). The scanning parameters included a source
voltage of 70 kV, tube current of 100 µA, image pixel size of 15 µm, exposure time
of 650 ms, and a rotation step of 0.2 to over 360 degrees.
Image reconstruction was performed using NRecon software version 1.7.3.1 (Bruker;
Kontich, Belgium) through Hamming-filtered back projection, with the following parameters:
smoothing (1), smoothing kernel (2—Gaussian), and ring artifact correction (5). µCT
images were resliced parallel to the axial plane, and the microarchitecture analysis
was performed on consecutive slices or with multiplanar image reconstructions. The
software rendered enamel structures in pink to facilitate identification.
Data Analysis
Amelogenin and FABP-3 expression levels were quantified using ImageJ for IHC-stained
sections and expressed in arbitrary units (AU). Tooth density was measured via micro-CT
imaging and reported in Hounsfield units (HU), allowing comparison of mineralization
between groups. All numerical data were analyzed using IBM SPSS Statistics 26. Data
are presented as mean values ± SD. The data showed normal distribution and homogeneous
variability (p > 0.05); thus, the variable means between the control and treated groups were compared
using the t-test. A p-value less than 0.05 (p < 0.05) was considered statistically significant.
Results
The immunohistochemical staining in [Figs 1] and [2] shows that the treatment group exhibits more intense staining, indicating higher
protein expression. Specifically, [Fig. 1] highlights pronounced amelogenin presence (red square) at 100× and 400× magnifications,
suggesting enhanced enamel matrix formation. Similarly, [Fig. 2] shows elevated FABP-3 expression, with more intense brown staining in the treatment
group at both magnifications.
 Fig. 1 Amelogenin expression (highlighted in red squares and indicated by red arrows) in
the tooth buds of fetal mice: (A) Control group at 100× and (B) 400× magnification; (C) treatment group receiving nanoparticle saltwater fish powder at 100× and (D) 400× magnification. The red arrows point to ameloblast cells exhibiting positive
amelogenin expression.
Fig. 1 Amelogenin expression (highlighted in red squares and indicated by red arrows) in
the tooth buds of fetal mice: (A) Control group at 100× and (B) 400× magnification; (C) treatment group receiving nanoparticle saltwater fish powder at 100× and (D) 400× magnification. The red arrows point to ameloblast cells exhibiting positive
amelogenin expression.
 Fig. 2 FABP-3 expression (red square) in fetal mice placental tissue: (A) control group at 100× and (B) 400× magnification; (C) treatment group receiving nanoparticle saltwater fish powder at 100× and (D) 400× magnification. More intense brown staining indicates higher FABP-3 expression
in the treatment group.
Fig. 2 FABP-3 expression (red square) in fetal mice placental tissue: (A) control group at 100× and (B) 400× magnification; (C) treatment group receiving nanoparticle saltwater fish powder at 100× and (D) 400× magnification. More intense brown staining indicates higher FABP-3 expression
in the treatment group.
The differences in staining intensity and color between the control and treatment
groups are likely due to variations in the level of amelogenin protein expression.
In immunohistochemical staining, higher protein expression typically results in darker
or more intense staining, whereas lower expression produces lighter staining. These
differences do not reflect inconsistent staining protocols, but somewhat true biological
variation in amelogenin expression between the groups. All slides were stained under
identical conditions to maintain consistency.
Quantitative analysis revealed significant differences between groups ([Table 1]). The mean amelogenin expression, measured as the mean staining intensity using
ImageJ analysis of IHC-stained sections, was 0.49 ± 0.40 AU in the control group and
increased significantly to 4.34 ± 3.26 AU in the treatment group (p = 0.005). FABP-3 expression was also significantly higher in the treatment group
(2.26 ± 0.85 AU) than in the control (1.50 ± 0.40 AU), with a p-value of 0.038.
Table 1
The numerical data of amelogenin and FABP-3 expression in control and treated groups
|
Variables
|
Control group
|
Treated group
|
p-Value
|
|
Amelogenin expression (AU)
|
0.49 ± 0.40
|
4.34 ± 3.26
|
0.005[a]
|
|
FABP-3 expression (AU)
|
1.50 ± 0.40
|
2.26 ± 0.85
|
0.038[a]
|
a A significant difference (p < 0.05) based on the T-independent test.
Additionally, μCT imaging displayed structural differences in enamel density between
groups. The enamel (pink) was more apparent and presumably denser in the treatment
group ([Fig. 3B]) compared to the control ([Fig. 3A]), with the red arrows indicating the area of interest.
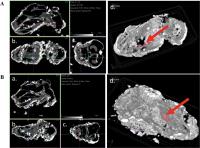 Fig. 3 Micro-computed tomography (μCT) scan of a fetal mouse head. The red arrow indicates
the region of interest (ROI), specifically the developing tooth germ area: (A) control group and (B) treated group—(a) sagittal section, (b) axial section, (c) anteroposterior section,
and (d) 3D-reconstruction. The enamel is rendered in pink.
Fig. 3 Micro-computed tomography (μCT) scan of a fetal mouse head. The red arrow indicates
the region of interest (ROI), specifically the developing tooth germ area: (A) control group and (B) treated group—(a) sagittal section, (b) axial section, (c) anteroposterior section,
and (d) 3D-reconstruction. The enamel is rendered in pink.
Quantitative analysis revealed a significant increase in enamel density in the treated
fetal mice (228.73 ± 5.31 HU) compared to the controls (220.75 ± 5.95 HU), with a
p-value of 0.022 (see [Table 2]).
Table 2
The numerical data of enamel density in control and treated groups
|
Variable
|
Control group
|
Treated group
|
p-Value
|
|
Enamel density (HU)
|
220.75 ± 5.95
|
228.73 ± 5.31
|
0.022[a]
|
a A significant difference (p < 0.05) based on the T-independent test.
Discussion
This study demonstrates that the administration of nanoparticle saltwater fish powder
significantly enhances amelogenin, FABP-3 expression, and enamel density during tooth
development. These findings suggest nanoparticle formulations improve bioavailability
and biological efficacy, positively influencing the key molecular pathways involved
in amelogenesis. The observed improvements in both protein expression and enamel density
underscore the therapeutic promise of this approach in supporting optimal dental tissue
formation.
Amelogenesis is a highly coordinated process involving multiple distinct stages characterized
by morphological and functional changes in ameloblasts.[3]
[17] The early presecretory stages involve odontoblasts-driven predentin, which initiates
mineralization at the dentinoenamel junction. Following dentin mineralization, preameloblasts
differentiate into secretory ameloblasts, which produce enamel matrix proteins, predominantly
amelogenin.[3]
[17]
[18] Amelogenin plays a crucial role in regulating mineral deposition and controlling
the morphology of enamel crystals.[7]
[19]
[20] In this study, immunohistochemical analysis revealed significantly increased amelogenin
expression in the treatment group receiving nanoparticle saltwater fish powder (4.34 ± 3.26)
compared to the control group (0.49 ± 0.40), with a p-value of 0.005. This upregulation indicates enhanced ameloblast activity and suggests
that the bioactive components within the nanoparticle formulation could stimulate
intracellular signaling pathways such as Wnt/β-catenin and BMP pathways, both of which
are well-documented regulators of ameloblast differentiation and enamel biomineralization.[21] The improved amelogenin expression likely contributed to the increased enamel density
observed via μCT.
Previous studies by Christiono et al using saltwater fish powder (non-nanoparticle)
reported a mean amelogenin expression of approximately 3.13.[6] The higher amelogenin expression observed in this study (4.34 ± 3.26) suggests that
nanoparticle delivery enhances the bioavailability of bioactive nutrients, facilitating
a more pronounced stimulation of ameloblast activity and protein expression during
the critical presecretory-to-secretory transition. This enhanced delivery aligns with
findings from nanotechnology research, which indicate that reducing particle size
to 50 to 300 nm improves cellular uptake and tissue penetration.[22]
Calcium plays a crucial role in the mineralization of enamel and dentin. The protein
calbindin-28kDa facilitates calcium diffusion and buffering within the ameloblast,
influencing the formation of nanospheres and the crystallization of hydroxyapatite.[23]
[24] Calcium ions can induce conformational changes in amelogenin, thereby enhancing
its self-assembly into nanospheres that regulate the growth of hydroxyapatite crystals.[25] The increased calcium content in the nanoparticle fish powder likely contributed
to higher calcium bioavailability, supporting enamel mineralization. Recent studies
have also highlighted innovative biomaterials that complement nutritional strategies
in promoting enamel remineralization. Dhivya et al reported that strontium-doped fluorophosphate
glasses exhibited enhanced ion release, improved bioactivity, and promoted apatite
formation, thereby demonstrating potential for effective surface enamel repair.[26] In parallel, Saravana Karthikeyan et al demonstrated that both synthetic and eggshell-derived
nanohydroxyapatite incorporated into carboxymethyl chitosan matrices facilitated biomimetic
mineral deposition with favorable cytocompatibility, highlighting their applicability
as sustainable and bioactive restorative agents.[27] Collectively, these findings suggest that ion-releasing glass systems and nanoparticle-based
composites provide complementary mechanisms of action, which may be further optimized
or integrated to achieve superior clinical outcomes in enamel remineralization.
Omega-3 fatty acids, including DHA, EPA, and alpha-linolenic acid (ALA), present in
saltwater fish, are known for their anti-inflammatory and osteogenic effects. These
fatty acids have been reported to promote calcium absorption, osteoblast differentiation,
and lipid oxidation, collectively supporting mineralization processes. FABPs are a
family of intracellular lipid-binding proteins that play a critical role in the uptake,
transport, and metabolism of long-chain fatty acids. Among the different isoforms,
FABP-3 (heart-type FABP) is particularly important in placental tissue due to its
high affinity for polyunsaturated fatty acids, including DHA and EPA. Transport of
omega-3 fatty acids to placental tissue is mediated by FABP-3.[28] Our observation of increased FABP-3 expression in the fetal trophoblast tissue indicates
facilitated transport of omega-3 fatty acids across the placenta, enhancing fetal
access to these nutrients.[29] By mediating the selective uptake and intracellular transport of these essential
fatty acids, FABP-3 ensures adequate fetal supply.
The anti-inflammatory properties of omega-3s may also protect ameloblasts from oxidative
stress, thereby maintaining their functional integrity during enamel formation.[19] Indahyani et al suggested that omega-3 supplementation affects tooth maturation
and overall mineralization quality by modulating cytokines and growth factors involved
in odontogenesis.[28] Similarly, Permatasari et al reported that omega-3 administration during pregnancy
in mice enhanced the mineral composition of enamel by increasing CaO and P2O5 levels.[30] Although direct human studies on enamel density are still limited, these findings
collectively support the hypothesis that omega-3 fatty acids contribute to enamel
development and mineralization.
Adding chitosan, a natural biopolymer derived from chitin, enhances membrane permeability
and facilitates the penetration of nanoparticles across biological barriers, including
the placenta.[16] Its properties, such as increased membrane contact and opening of tight junctions,
allow for more efficient delivery of calcium, phosphorus, and omega-3 fatty acids
to fetal tissues, including ameloblasts. Improved nutrient transport correlates with
increased ionic availability, promoting mineralization and matrix organization, reflected
in the higher enamel density observed via μCT analysis.[21]
[31]
The findings of this study suggest that nanoparticle-based nutrient delivery systems
hold significant potential for prenatal interventions to optimize dental enamel development.
Such strategies could be particularly beneficial in populations vulnerable to mineral
deficiencies or developmental enamel defects, such as amelogenesis imperfecta or fluorosis.
Further research should investigate the detailed molecular signaling pathways involved,
including those regulating Wnt/β-catenin, BMP, and other pathways critical to tooth
morphogenesis. Longitudinal studies are warranted to investigate the durability and
functional integrity of the newly formed enamel. Additionally, examining the effects
of other bioactive compounds incorporated into nanoparticle formulations could expand
the therapeutic spectrum. Translational research should also assess the safety profile
and optimal dosing of nanoparticle nutrients to ensure efficacy without adverse effects.
This study was conducted during the pre-secretory stage; therefore, the findings only
reflect early amelogenin expression and may not fully represent its functional role
in the later stages of enamel development.
Conclusion
Nanoparticle saltwater fish powder demonstrates promising potential in enhancing amelogenin
and FABP-3 expression, promoting mineralization, and increasing enamel density during
fetal development. These findings underscore the importance of integrating nanotechnology
with nutrient supplementation strategies to improve prenatal dental health outcomes.





