

Subscribe to RSS
DOI: 10.1055/s-0043-1778013
Alcohol and Caffeine Co-Administration Increased Acetylcholinesterase Activity and Inflammatory Cytokines in Sleep-Deprived Rats: Implications for Cognitive Decline and Depressive-Like Manifestations
Authors
Funding The authors declare that they have not received funding form agencies in the public, private, or non-profit sectors for the conduction of the present study.

Abstract
Sleep deprivation is a major health problem in modern society; it has been worsened by alcohol and caffeine intake to stay awake and improve bodily activities, an experience common among night-shift workers. For the present study, 50 adult male Wistar rats weighing between 150 g and 200 g were randomly selected and divided into 5 groups of 10 rats each (n = 10). Group 1 was the control group; group 2 was the group of sleep-deprived (SD) rats; group 3 was composed SD rats submitted to the administration of 20% alcohol; group 4 comprised SD rats submitted to the administration of 200 mg/kg of caffeine; and Group 5 was composed of SD rats who underwent the co-administration of 20% alcohol and 200 mg/kg of caffeine. At the end of 28 days, the animals were euthanized, and blood samples were collected for biochemical analysis. Memory, anxiety, social behavior and locomotive activity were assessed using the Y-maze, the elevated plus maze, the hole-board and three-chambered social approach tests, and the open field test. The plasma levels of the acetylcholinesterase (AChE) enzyme and inflammatory cytokines (interleukin 6 [IL-6], interleukin 10 [IL-10], and tumor necrosis factor beta, [TNF-β]) were also measured. Data was expressed as mean ± standard error of the mean [SEM] values, and the data were analyzed through analysis of variance (ANOVA) followed by the Tukey post hoc test, with significance set at p < 0.05. The results revealed that sleep deprivation, and the co-administration of alcohol and caffeine impair memory in rats. Sleep deprivation also caused a significant increase in anxiety and anxiety-related behavior, with decreased social interaction, in rats. Locomotive activity was improved in SD rats, especially in those to which alcohol was administered. Sleep deprivation significantly reduced acetylcholinesterase activity among SD rats and those to which alcohol was administered when compared with the controls. The plasma levels of IL-6, IL-10 and TNF-β were significantly increased in SD rats when compared with the controls. The administration of alcohol and caffeine separately, as well as their co-administration, significantly increased cytokine levels in rats.
Keywords
sleep deprivation - alcohol - caffeine - inflammatory cytokines - acetylcholinesterase activityIntroduction
Alcohol, which is addictive to humans, is a psychotropic substance found in beer, wine, and spirits, and the fifth leading cause of cancer, with effects ranging from impairment of neurocognitive function, dizziness, nausea, vomiting, and hangover-like symptoms to liver and brain damage.[1] [2] [3] [4] Caffeine, on the other hand, is a methylxanthine central nervous system (CNS) stimulant, used as a cognitive enhancer to increase alertness and attentional performance, physical stamina, mood, memory, and cognitive performance.[5] [6] [7] [8] [9] [10] At moderate amounts, caffeine can stimulate behavior which may indicate CNS activation, but, at high concentrations, it can even decrease learning and memory-related behavior.[11] [12] Pharmacologically, caffeine is an adenosine-receptor antagonist that primarily affects the A1 and A2A receptors, which are connected to brain activities related to sleep, arousal, and cognition; it takes effect within the first 30 minutes after intake due to its rapid absorption in the stomach and small intestine.[13] However, the effects of caffeine overdose range from tachycardia, hypertension, nausea, vomiting, gastritis, irritation, sleeplessness, sadness, anxiety, and seizures, to stroke.[14] A major reason for the deliberate consumption of caffeine is to combat sleepiness.[11]
Sleep deprivation has become a global public health problem associated with various social consequences, such as obesity, type-2 diabetes, hypertension, cardiovascular disease, anxiety, and depression.[15] [16] [17] [18] Sleep deprivation impairs cognitive function and reduces cognitive control, and it causes disruptions to capacity planning, ethical misbehavior, and anxiety.[19] [20] [21] Sleep deprivation impairs acetylcholinesterase activity, thus causing decreased cholinergic release and/or via glutamatergic inhibition.[22] Insufficient sleep is one of the most prevalent and important health problems worldwide, and it is associated with immune system modulation and mood and immune system declines.[23] [24] Due to the pressures of the workplace, the growth of urban centers, modern civilization, societal factors such as cost of living, safety of the environment in which one lives, increased incidence of anxiety, depression, and other psychiatric disorders, individuals are plagued by sleep deprivation, which raises the levels of inflammatory biomarkers, thus affecting their health and well-being.[25] Sleep deprivation impairs immune function, leading to alterations in inflammatory cytokines.[26] [27]
Inflammatory biomarkers are used to monitor chronic disease activity and overall health status; thus, they act as useful indicators of infection.[28] [29] Cytokines, which are involved in cell signaling, are the actual components that signal the immune system to respond in a specific area of the body that is being invaded.[30] When cytokines are elevated, this indicates that the body's immune system has been triggered and there is inflammation occurring somewhere in the body.[31] Anti-inflammatory cytokines (such as interleukin 6 [IL-6] and interleukin 10 [ IL-10]) regulate the production of pro-inflammatory cytokines, hence blocking the synthesis of IL-1, tumor necrosis factor (TNF), and other important pro-inflammatory cytokines. Interleukin 6 reduces inflammation though the inhibition of TNF and IL-1 production by macrophages and stimulation of acute-phase proteins, plasma proteins whose concentrations change during acute inflammatory responses.[32] [33] Transforming growth factor-beta (TGF-β) often exhibits disparate effects, with immune-enhancing activity in local tissues and immune-suppressive activity in the systemic circulation, by suppressing the proliferation and differentiation of T cells and B cells, and it limits the production of IL-2, interferon-gamma (IFN-γ), and TNF.[32]
It is well established that sleep plays a critical role in learning and memory formation. Studies have demonstrated that sleep deprivation in animals leads to memory deficits in several behavioral models.[34] It has been proposed that the deleterious effects caused by sleep deprivation are due to its long-term effect in the hippocampus, which is important for memory.[35] The mechanisms by which altered sleep duration affects health are unclear, but experimental studies[36] [37] [38] [39] suggest that altered sleep may impact levels of cytokines known to be important in regulating inflammation. These studies[36] [37] [38] [39] show that sleep deprivation results in various medical conditions, such as cardiovascular disease, arthritis, diabetes mellitus, altered-immune responses in certain cancers, and increased activation of cellular signals that initiates the expression of inflammatory cytokines. Alcohol affects virtually every system, including the immune system, and heavy alcoholism is associated with increased cytokine secretion and inflammation in these organs, reinforcing the possibility that cytokines play a pivotal role in alcohol-related brain and heart damage.[40] [41] It becomes pertinent for the present study to investigate the result of altered cognitive decline and depressive-like manifestations implicated in increased acetylcholinesterase activity and inflammatory cytokines in sleep-deprived (SD) rats.
Materials and Methods
Experimental Animals and Grouping
In total, 50 adult male Wistar rats weighing between 150 g and 200 g were obtained and housed in the animal unit of the Faculty of Basic Medical Sciences, Delta State University, Abraka, under standard laboratory conditions and grouped into 5 groups of 10 rats (n = 10) as follows:
-
Group 1: Control;
-
Group 2: SD Wistar rats;
-
Group 3: SD + 20% alcohol;
-
Group 4: SD + 200 mg/kg of caffeine; and
-
Group 5: SD + 20% alcohol + 200 mg/kg of caffeine.
Induction of Sleep Deprivation
Sleep deprivation was induced using the modified multiple platform model as described by Nunes and Tufik (1994). In this method, the animals are kept for 8 hours a day on 12 small circular (3 cm in diameter) platforms each that are 41 cm high inside a chamber that measuring 23 × 234 × 35 cm and filled with water up to 1cm below the upper surface. The animals are returned to their respective cages after the daily 8 hours of sleep deprivation. This process continues for four weeks. The animals lose muscle tone at the start of each episode of paradoxical sleep, which causes them to fall into the water and awaken.
Alcohol and Caffeine Reconstitution and Administration
For administration of alcohol (20%), 20 mL of ethanol were diluted in 80 mL of distilled water, and 400 mg of caffeine were diluted in 10 mL of normal saline to obtain a stock solution of 40 mg/mL. A dose of 200 mg/kg was then administered based on the rat's body weight. Alcohol and caffeine were administered by orogastric gavage once a day between 8am and 10am for a period of 4 weeks.
Behavioral Studies
Working memory, anxiety, social behavior, and locomotive activity were assessed.[42] [43] [44] Working memory performance was assessed by recording spontaneous alternation behavior in a single session in a modified Y-maze model with 3 equal arms at an angle of 120° from each other. The hole-board test is a mice apparatus that was adapted for the dimensions of a rat. Each rat was placed at the end of one arm and allowed to move freely through the maze during an 8-minute session. The series of arm entries were recorded visually. Entry was considered completed when the hind paws of the rat had completely entered the arm. Alternation was defined as successive entries into three different arms (A, B, and C) of overlapping triplet sets.[45] Percentage alternation was calculated as the ratio of actual to possible alternation (defined as the total number of arm entries minus 2), multiplied by 100, as follows: % alternation = (number of alternations)/(Total arm entries - 2) x 100.
Anxiety was assessed using the hole-board test and elevated plus maze (EPM) test, as described by a previous studies.[46] The EPM apparatus consists of 2 open arms (measuring 25 × 5 cm) and 2 closed arms (measuring 25 × 5 × 16 cm) arranged perpendicularly and connected at the center (measuring 5 × 5 cm) and elevated to a height of 100 cm. Each rat was placed in the center of the apparatus, and the number of entries and time spent per open and closed arm, respectively, were recorded. The hole-board consists of a square board with 16 holes, each measuring around 3 cm in diameter.[42] However, it was modified to 6 cm for rats in the current study to measure specific animal behavior by head dip, which is considered a measure of neophilia.[47] The anxiety level is inversely proportional to the number of head dips.[48]
Social behavior was assessed through the three-chambered social approach test, as described by a previous study,[49] which is based on two behaviors: social avoidance and social fear. In this test, each animal is placed in one of the chambers and allowed to explore it for five minutes. Thereafter, an unfamiliar animal is placed in another chamber. The time the test animal spends alone in its chamber and the time the animal spends interacting with the unfamiliar (stranger) animal is observed and recorded. Social avoidance is considered as a decrease in the duration of the interaction, and social fear is shown by behaviors such as flight, defensive burying, and alarm cries.
Locomotive activity was assessed through the open field test, as described by previous research.[50] The open field used in the present study is a rectangular wooden arena composed of a floor measuring 36 × 36 × 26 cm. The floor was divided with permanent red markings into 16 equal squares at the bottom. It was placed in a sound-isolation room with dim lighting. Locomotion (number of floor units entered with all paws), rearing frequency (number of times the animal stood on its hind legs or with its forearms against the walls of the observation cage or free in the air), and frequency of grooming (number of body cleanings with paws, picking of the body and pubis with the mouth, and face-washing actions) were recorded for thirty minutes and scored. The trial lasted for 5 minutes. An increase in locomotion, rearing, and grooming connotes a central excitatory response, while a decrease connotes central inhibition.
Sample Collection
At the end of the four weeks, the rats fasted overnight and were euthanized by cervical dislocation, and laparotomy was performed to open the animals. A blood sample was collected from the inferior vena cava and placed inside a labeled plain container to obtain serum for the biochemical analysis.
Enzyme Assay
After decapitation, the brains were excised and kept on a Petri dish cooled with crushed ice. The brains were washed with cold isotonic saline to remove blood, and the brain regions were then immediately dissected and weighed. The tissue was kept in cold 0.32 M sucrose (pH 7.4 with Tris base). Homogenates (5% w/v) were prepared using glass homogenizer tubes and a Teflon (The Chemours Company, Wilmington, DE, United States) motor-driven pestle. The homogenates were centrifuged at 900 g for 10 minutes at 4°C, and the supernatants were collected and centrifuged at 100,000 g for 60 minutes at 4°C.
Biochemical Analysis
Lipid Peroxidation Assay
Lipid peroxidation was determined as the formation of malondialdehyde (MDA) according to the method of Ohkawa et al.[51]
Assessment of the Activity of Acetylcholinesterase (AChE)
Acetylcholinesterase (AChE) activity was determined by the method of Ellman et al.[52]
Estimation of Cytokine Level
Plasma samples (100 μl; 1:50 dilution in assay buffer) were assayed for ILs (IL-6 and IL-10) and TNF-β using commercial enzyme-linked immunosorbent assay (ELISA) kits. The assays were performed as per the manufacturer's protocols.
Ethical Consideration
Ethical approval (number: RBC/FBMS/DELSU/P22/013) was obtained from the Ethics in Research Committee of the Faculty of Basic Medical Sciences, Delta State University, Abraka, Nigeria, for the use of laboratory animals.
Statistical Analysis
The results were expressed as mean ± standard error of the mean (SEM) values. The statistical comparisons were performed using one-way analysis of variance (ANOVA), followed by the Tukey post hoc test for multiple comparisons, using the Prism statistical software (GraphPad Software, La Jolla, CA, United States), version 9), and values of p ≤ 0.05 were considered statistically significant.
Results
[Figure 1] shows the assessment of memory using the Y-maze test in SD male Wistar rats; sleep deprivation significantly (p < 0.05) impaired memory function in all groups 3, 4, and 5 (SD rats which underwent the administration of alcohol, caffeine, and both respectively) when compared with the controls. Similarly, significant (p < 0.05) memory impairments were observed in groups 3, 4, and 5 when compared to group 2 (SD rats not submitted to the administration of caffeine or alcohol). The lowest level of memory impairment was observed in group 5 when compared to groups 3 and 4.

[Figure 2] shows the impact of the administration of caffeine and alcohol on anxiety-related behavior using the EPM; compared to the controls, sleep deprivation caused a significant (p < 0.05) increase in the level of anxiety of groups 3, 4, and 5. There was also a significant (p < 0.05) increase in the level of anxiety of groups 3, 4, and 5 compared to group 2. The anxiety level among group 5 was significantly (p < 0.05) higher when compared to that of group 4.
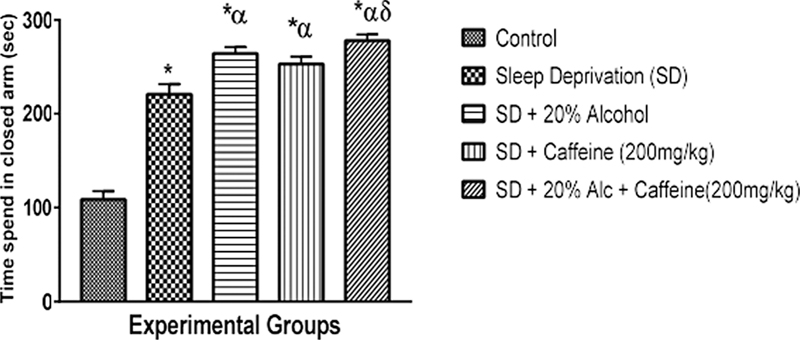
[Figure 3] shows the assessment of the anxiety level using the hole-board test. Sleep deprivation significantly (p < 0.05) increased the anxiety levels of groups 3, 4, and 5 when compared to the controls. However, this increase was higher in group 3. The data also shows that the anxiety level significantly (p < 0.05) decreased in group 5 when compared to groups 2 and 3. Similarly, the level of anxiety in group 4 significantly decreased when compared to group 3.

[Figure 4] shows the assessment of social behavior using the three-chambered social approach test. Sleep deprivation caused a significant (p < 0.05) decrease in social behavior in groups 3, 4, and 5 when compared to the controls. The lowest level of decrease was observed in group 5. Social behavior also significantly (p < 0.05) decreased in groups 3, 4, and 5 compared to group 2.

[Figure 5] shows the assessment of motor activity using the open field test. Sleep deprivation significantly (p < 0.05) increased motor activity in groups 2, 3, and 4 compared to the controls; this increase was highest in group 3. However, a significant (p < 0.05) decrease in motor activity was observed in group 5 compared to the controls and to group 2.

[Figure 6] shows the assessment of acetylcholinesterase activity. A significant (p < 0.05) decrease in acetylcholinesterase activity was observed in groups 2, 3, and 4 compared to the controls; and a significant (p < 0.05) increase was observed in group 3 compared to the controls. Inversely, acetylcholinesterase activity was significantly (p < 0.05) reduced in group 4 compared to group 2; there was also a significant (p < 0.05) reduction in group 5 compared to the controls and group 2.

[Figure 7] shows the assessment of the levels of TNF-β. Sleep deprivation significantly (p < 0.05) increased the levels of TNF-β in groups 3, 4, and 5 compared to the controls, but the difference in the increase in groups 3, 4, and 5 compared to group 2 was not significant. Comparisons involving only groups 3, 4, and 5 did not show significant differences either.

[Figure 8] presents the assessment of IL-6 levels. Sleep deprivation significantly (p < 0.05) increased the levels of IL-6 in of groups 3, 4, and 5 compared with the controls. A significant (p < 0.05) increase was also observed in group 4 compared to group 2.

[Figure 9] presents the assessment of IL-10 levels. Sleep deprivation significantly (p < 0.05) increased the levels of IL-10 in groups 3, 4, and 5 compared to the controls. A significant (p < 0.05) increase was also observed in group 4 compared to group 5. And a slightly significant (p < 0.05) decrease was observed in group 3 compared to group2.

Discussion
The results of the present study revealed that sleep deprivation significantly impaired memory in groups 3, 4, and 5 compared with the controls ([Fig. 1]). This effect may be attributed to impairment of the hippocampus through the destruction of synaptic plasticity and a decrease in the speed of processing information during sleep deprivation. However, we must also considering that alcohol affects memory by altering hippocampal cell-to-cell communication, which plays an important role in forming and maintaining memory.[53] Our finding was consistent with the study by Vedder et al,.[54] who also reported the association of cognitive dysfunction and excessive consumption of alcohol. In the present study, memory impairment was observed in SD rats that received only caffeine when compared to the controls and non-treated SD rats. This is in line with the study by Chen et al.,[55] who reported that sleep deprivation alters the beneficial effects on the brain of caffeine, a known CNS stimulant which can alleviate brain fatigue and low cognitive efficiency. The mechanism by which caffeine brings about wakefulness is by blocking sleep-promoting receptors in the brain called adenosine receptors, which bind to adenosine, thus blocking it and preventing its activation.
In the current study, sleep deprivation caused anxiety in groups 3, 4, and 5 compared to the controls using the Y-maze test, EPM, and the hole-board test ([Fig. 2] [3] [4]). The decrease in anxiety when using hole-board test was observed in the comparison of group 5 to groups 2 and 3, while the Y-maze test showed the same effect when compared to the controls but the opposite effect when compared to group 2. This is in line with the study by Chang et al.,[56] who reported that sleep deprivation significantly caused anxiety. In the current study, in the EPM and hole-board test models, the level of anxiety significantly increased in the groups who had received alcohol (groups 3 and 5) when compared to control group. This may be due the potential of alcohol to make anxiety progressively worse, as it is a CNS depressant that alters the levels of serotonin and other neurotransmitters in the brain, thus interfering with the fight-or-flight response to cause anxiety.[57] [58] [59]
Furthermore, a significant increase in the level of anxiety was also observed in group 4 when compared group 2. The level of anxiety in group 5 also significantly increased. These results are in line with those of a previous study[60] that reported the effect of caffeinated drinks on sleep. The authors[60] hypothesized that kids aged 6 to 10 years who regularly consume an average of 0.4 mg of caffeinated sleep for about 15 minutes. Another study[61] reported that the sleep of subjects who drink 100 mg to 150 mg of caffeine a day was more interrupted than that of subjects who drink lower amounts, and that sleep disturbances were worse on nights after higher consumption, because the effects of alcohol may be increased by caffeine's inhibition of A2A receptors.
Data from the present study revealed that sleep deprivation caused a significant (p < 0.05) decrease in social behavior in groups 3, 4, and 5 compared to the controls ([Fig. 4]). Such an alteration may be linked to the reports of previous studies that chronic sleep deprivation, for longer than a week, causes changes in the neurotransmitter receptor systems and neuroendocrine stress systems; these alterations match those linked to severe depression.[62] Moreover, excessive drinking reduces serotonin, and low serotonin levels result in depression.[63] Our findings were contrary to the findings of Al-Maddah et al.[64] that chronic rather than acute sleep deprivation may result in depression.
Prolonged sleep deprivation and the administration of alcohol and caffeine alter motor activity ([Fig. 5]). This finding was similar to that of a previous study[65] that reported no significant increase in motor activity under 24 hours of oxidative stress. However, another study[66] reported a finding to the contrary: reduced motor activity in sleep deprivation, including the ability to execute instructions. The administration of alcohol to SD rats significantly increased motor activity when compared to non-treated sleep deprived rats in the current study. This observation correlates to that of a previous study,[65] which reported that the brain adenosine system represents a common pathway for the effects of ethanol and sleep deprivation on motor activity. Caffeine-treated rats were observed to display increased motor activity when compared to the control group. This observation may be attributed to the antagonist measure of caffeine against poor motor activity performance in sleep deprivation by blocking receptor binding and thus has different neurochemicals effects.[64] [65] However, the co-administration of alcohol and caffeine in SD rats significantly decreased motor activity. Such a decrease could be due to the oxidative stress caused by the co-administration of alcohol and caffeine potentiating differential neurochemical effects.[67]
Among its various effects, sleep deprivation has been shown to decrease AChE activity in rats, as observed in the current study ([Fig. 6]), an effect which can lead to the accumulation of AChE at the synaptic junctions, causing cytotoxicity. Our results support the earlier findings of Vidyasagar et al.,[68] who reported that 96 hours of sleep deprivation significantly reduced whole-body AChE activity, which is suggestive of high acetylcholine turnover in this region. In the present study, the administration of 20% alcohol amid sleep deprivation significantly increased the level of AChE activity, which results in decreased cholinergic activity, following increased AChE activity and vice versa. This is in line with previous studies[22] that reported that alcohol increases AChE activity in SD Wistar rats. Similarly, our findings corroborates that of previous studies:[69] [70] caffeine significantly decreases AChE activity, which was shown by a significantly increased level of acetylcholine in the brain. Meanwhile, caffeine has been shown to delay or prevent sleep, improving task performance during sleep deprivation by binding to antagonizing adenosine receptors in the brain.[71]
In the present study, we observed significantly higher levels of circulating TNF-β, IL-6, and IL-10 in the serum of both SD rats and the SD rats treated with alcohol and caffeine when compared to the levels of the controls ([Figs. 7] [8] [9]). Tumor necrosis factor beta is involved in the regulation of cell survival, proliferation, differentiation, and apoptosis; it also inhibits tumor growth and kills malignant cell lines, and plays a significant role in innate immune control.[72] However, circulating inflammatory cytokines have been identified to be accelerated in alcohol and caffeine-induced patients and several inflammatory cytokines including IL-6, IL-10 and TNF-β. This shows that the CNS is impacted by increased quantities of circulating cytokines, resulting in altered sleep-wake cycle or troubled sleep.[73] [74] [75] This observation corroborates the findings of the present study, despite the fact that our sample was different from the samples of these other studies.[73] [74] [75] Data from the present study has also shown that the levels of inflammatory biomarker TNF-β were elevated in SD rats. This corroborates previous findings[76] [77] [78] [79] that a high percentage of people with anxiety-related disorders caused by sleep deprivation exhibit evidence of elevated inflammatory markers in blood concentrations and their soluble receptors.
Conclusion
Sleep deprivation and the co-administration of alcohol and caffeine impaired the memory of the experimental rats compared to control rats. Sleep deprivation further caused a significant increase in anxiety and anxiety-related behavior, with decreased social interaction. Locomotive activity was improved in SD rats, especially in those treated with alcohol. Sleep deprivation significantly reduced AChE activity among SD rats and SD rats treated with alcohol when compared with the controls. The isolated and combined administration of alcohol and caffeine significantly increased the levels of these cytokines in rats. Overall, the co-administration of alcohol and caffeine increased AChE activity and the levels of inflammatory cytokines in SD rats with possible cognitive decline and depressive-like manifestations.
Conflict of Interests
The authors have no conflict of interests to declare.
Consent for Publication
Not Applicable.
Availability of Data and Material
The data of the present study may be made available based on reasonable request to the corresponding author.
-
References
- 1 Kamath J, Virdi S, Winokur A. Sleep disturbances in schizophrenia. Psychiatr Clin North Am 2015; 38 (04) 777-792
- 2 Feeley CA, Turner-Henson A, Christian BJ. et al. Sleep quality, stress, caregiver burden, and quality of life in maternal caregivers of young children with bronchopulmonary dysplasia. J Pediatr Nurs 2014; 29 (01) 29-38
- 3 Thakkar MM, Sharma R, Sahota P. Alcohol disrupts sleep homeostasis. Alcohol 2015; 49 (04) 299-310
- 4 Younes M, Hanly PJ. Immediate postarousal sleep dynamics: an important determinant of sleep stability in obstructive sleep apnea. J Appl Physiol 2016; 120 (07) 801-808
- 5 Clark I, Landolt HP. Coffee, caffeine, and sleep: A systematic review of epidemiological studies and randomized controlled trials. Sleep Med Rev 2017; 31: 70-78
- 6 Cappuccio FP, D'Elia L, Strazzullo P, Miller MA. Quantity and quality of sleep and incidence of type 2 diabetes: a systematic review and meta-analysis. Diabetes Care 2010; 33 (02) 414-420
- 7 McLellan TM, Caldwell JA, Lieberman HR. A review of caffeine's effects on cognitive, physical and occupational performance. Neurosci Biobehav Rev 2016; 71: 294-312
- 8 Southward K, Rutherfurd-Markwick KJ, Ali A. The effect of acute caffeine ingestion on endurance performance: a systematic review and meta- analysis. Sports Med 2018; 48 (08) 1913-1928
- 9 Borota D, Murray E, Keceli G. et al. Post-study caffeine administration enhances memory consolidation in humans. Nat Neurosci 2014; 17 (02) 201-203
- 10 Sane RM, Jadhav PR, Subhedar SN. The acute effects of decaffeinated versus caffeinated coffee on reaction time, mood and skeletal muscle strength. J Basic Clin Physiol Pharmacol 2019; 30 (05) 12
- 11 Watson EJ, Coates AM, Kohler M, Banks S. Caffeine consumption and sleep quality in Australian adults. Nutrients 2016; 8 (08) 11-17
- 12 Howell S, Reid JL, Hammond D, McCrory C, Dubin JA, Leatherdale ST. Use of caffeinated energy drinks among secondary school students in Ontario: Prevalence and correlates of using energy drinks and mixing with alcohol. Can J Public Health 2017; 25: 106
- 13 Einöther SJ, Giesbrecht T. Caffeine as an attention enhancer: reviewing existing assumptions. Psychopharmacology (Berl) 2013; 225 (02) 251-274
- 14 Kang S, Yoo H, Chung S, Kim C. Eveningness, sleep patterns, daytime sleepiness and fatigue in Korean male adolescents. Sleep Med Psychophysiol 2012; 19: 89-96
- 15 Boivin DB, Boudreau P. Impacts of shift work on sleep and circadian rhythms. Pathol Biol (Paris) 2014; 62 (05) 292-301
- 16 Meerlo P, Havekes R, Steiger A. Chronically restricted or disrupted sleep as a causal factor in the development of depression. Curr Top Behav Neurosci 2015; 25: 459-481
- 17 Malish S, Arastu F, O'Brien LM. A preliminary study of new parents, sleep disruption, and driving: a population at risk?. Matern Child Health J 2016; 20 (02) 290-297
- 18 Ryu HS, Lee SA, Lee GH, Chung YS, Kim WS. Subjective apnoea symptoms are associated with daytime sleepiness in patients with moderate and severe obstructive sleep apnoea: a retrospective study. Clin Otolaryngol 2016; 41 (04) 395-401
- 19 Welsh T, Mai K, Ellis A, Christian M. Overcoming the effects of sleep deprivation on unethical behavior: an extension of integrated self-control theory. J Exp Soc Psychol 2018; 76: 142-154
- 20 Slama H, Chylinski D, Deliens G, Leproult R, Schmitz R, Peigneux P. Sleep deprivation triggers cognitive control impairments in task-goal switching. Sleep Epub. 2017; 14: 33-39
- 21 Barner C, Altgassen M, Born J, Diekelmann S. Effects of sleep on the realization of complex plans. J Sleep Res 2019; 28 (01) e12655
- 22 Thakkar M, Mallick BN. Effect of REM sleep deprivation on rat brain acetylcholinesterase. Pharmacol Biochem Behav 1991; 39 (01) 211-214
- 23 Paul W, John MH, Brieann CS, Devon AG, Kimberly AH, Hans VD. Sleep Deprivation diminishes attentional control effectiveness and impairs flexible adaptation to changing conditions. Sci Rep 2017; 7 (01) 1-9
- 24 Krause AJ, Simon EB, Mander BA. et al. The sleep-deprived human brain. Nat Rev Neurosci 2017; 18 (07) 404-418
- 25 Zhao SZ, Wang MP, Viswanath K. et al. Short sleep duration and insomnia symptoms were associated with lower happiness levels in Chinese adults. Int J Environ Res Public Health 2019; 16 (12) 2079
- 26 Ali T, Choe J, Awab A, Wagener TL, Orr WC. Sleep, immunity and inflammation in gastrointestinal disorders. World J Gastroenterol 2013; 19 (48) 9231-9239
- 27 Hurtado-Alvarado G, Domínguez-Salazar E, Pavon L, Velázquez-Moctezuma J, Gómez-González B. Blood brain barrier disruption induced by chronic sleep loss: low-grade inflammation may be the link. J Immunol Res 2016; 2016: 4576012
- 28 Wells G, Becker JC, Teng J. et al. Validation of the 28-joint Disease Activity Score (DAS28) and European League Against Rheumatism response criteria based on C-reactive protein against disease progression in patients with rheumatoid arthritis, and comparison with the DAS28 based on erythrocyte sedimentation rate. Ann Rheum Dis 2009; 68 (06) 954-960
- 29 Rawat R, Humphrey JH, Mutasa K, Ntozini R, Stoltzfus RJ. Short communication: predicting adverse HIV-related outcomes in a resource-limited setting: use of the inflammation marker α(1)-acid glycoprotein. AIDS Res Hum Retroviruses 2010; 26 (11) 1171-1174
- 30 Neurath F. Mesenchymal stem cells for chronic wound healing. Nat Rev Immunol 2014; 14: 329
- 31 Ogbutor UG, Nwangwa EK, Nwogueze BC. et al. Proinflammatory and Anti-inflammatory Cytokine Response to Isometric Handgrip Exercise and the Effects of Duration and Intensity of the Isometric Efforts in Prehypertensive Participants. J Chiropr Med 2022; 21 (03) 177-186
- 32 Barnes TC, Anderson ME, Moots RJ. The many faces of interleukin-6: the role of IL-6 in inflammation, vasculopathy, and fibrosis in systemic sclerosis. Int J Rheumatol 2011; 2011: 721608
- 33 Gabay C. Interleukin-6 and chronic inflammation. Arthritis Res Ther 2006; 8 (Suppl. 02) S3
- 34 Javad-Moosavi BZ, Nasehi M, Vaseghi S, Jamaldini SH, Zarrindast MR. Activation and inactivation of nicotinic receptors in the dorsal hippocampal region restored negative effects of total (TSD) and REM sleep deprivation (RSD) on memory acquisition, locomotor activity and pain perception. Neuroscience 2020; 433: 200-211
- 35 Rezaie M, Nasehi M, Vaseghi S. et al. The interaction effect of sleep deprivation and cannabinoid type 1 receptor in the CA1 hippocampal region on passive avoidance memory, depressive-like behavior and locomotor activity in rats. Behav Brain Res 2021; 396: 112901
- 36 Irwin MR, Opp MR, Mark RO. Sleep Health: Reciprocal Regulation of Sleep and Innate Immunity. Neuropsychopharmacology 2017; 42 (01) 129-155
- 37 Ferrucci L, Fabbri E, Sarwan N. Inflammageing: chronic inflammation in ageing, cardiovascular disease, and frailty. Nat Rev Cardiol 2018; 15 (09) 505-522
- 38 Shearer WT, Reuben JM, Mullington JM. et al. Soluble TNF-alpha receptor 1 and IL-6 plasma levels in humans subjected to the sleep deprivation model of spaceflight. J Allergy Clin Immunol 2001; 107 (01) 165-170
- 39 Andreakos E, Bierhaus A, Akerstedt T. Short Communication: Sleep loss activates cellular markers of inflammation. Brain Behav Immun 2017; 24 (01) 54-57
- 40 Fernández-Solà J, Planavila P. Cardiovascular risks and benefits of moderate and heavy alcohol consumption. Nat Rev Cardiol 2015; 12 (10) 576-587
- 41 Bishehsari F, Magno E, Swanson G. et al. Alcohol and Gut-Derived Inflammation. Alcohol Res 2017; 38 (02) 163-171
- 42 Desingu R, Balakrishnan S, Shamsher SK, Bhawna L. Animal models of anxiety: a review. Int J Basic Clin Pharmacol 2023; 12 (01) 134-141
- 43 Habibur R, Muralidharan P, Sivaraman D, Dipankar S. Continuous sleep deprivation for 5 days produces loss of memory in mice and may be a cause of Alzheimer's disease. Ann Biol Res 2010; 1 (04) 185-193
- 44 Silva RH, Kameda SR, Carvalho RC. et al. Anxiogenic effect of sleep deprivation in the elevated plus-maze test in mice. Psychopharmacology (Berl) 2004; 176 (02) 115-122
- 45 Sarter M, Bodewitz G, Stephens DN. Attenuation of scopolamine-induced impairment of spontaneous alteration behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology (Berl) 1988; 94 (04) 491-495
- 46 Komada M, Takao K, Miyakawa T. Elevated plus maze for mice. J Vis Exp 2008; (22) 1088
- 47 Hart PC, Bergner CL, Smolinsky AN. et al. Experimental Models of Anxiety for Drug Discovery and Brain Research. In: Proetzel G, Wiles MV. eds. Mouse Models for Drug Discovery. New York, NY: Springer New York; 2016: 271-291
- 48 Campos AC, Fogaça MV, Aguiar DC, Guimarães FS. Animal models of anxiety disorders and stress. Br J Psychiatry 2013; 35 (Suppl. 02) S101-S111
- 49 Toth I, Neumann ID. Animal models of social avoidance and social fear. Cell Tissue Res 2013; 354 (01) 107-118
- 50 Onaolapo OJ, Onaolapo AY, Akanmu MA, Olayiwola G. Foraging enrichment modulates open field response to monosodium glutamate in mice. Ann Neurosci 2015; 22 (03) 162-170
- 51 Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 1979; 95 (02) 351-358
- 52 Ellman GL, Courtney KD, Andres Jr V, Feather-Stone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 1961; 7: 88-95
- 53 Peng Z, Dai C, Ba Y, Zhang L, Shao Y, Tian J. Effect of Sleep Deprivation on the Working Memory-Related N2-P3 Components of the Event-Related Potential Waveform. Front Neurosci 2020; 14: 469
- 54 Vedder LC, Hall JM, Jabrouin KR, Savage LM. Interactions between chronic ethanol consumption and thiamine deficiency on neural plasticity, spatial memory, and cognitive flexibility. Alcohol Clin Exp Res 2015; 39 (11) 2143-2153
- 55 Chen X, Zhang L, Yang D. et al. Effects of Caffeine on Event-Related Potentials and Neuropsychological Indices After Sleep Deprivation. Front Behav Neurosci 2020; 14: 108
- 56 Chang YS, Chen HL, Wu YH, Hsu CY, Liu CK, Hsu C. Rotating night shifts too quickly may cause anxiety and decreased attentional performance, and impact prolactin levels during the subsequent day: a case control study. BMC Psychiatry 2014; 14 (01) 218
- 57 Holmes A, Fitzgerald PJ, MacPherson KP. et al. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-mediated fear extinction encoding. Nat Neurosci 2012; 15 (10) 1359-1361
- 58 Anker JJ, Kushner MG. Co-Occurring Alcohol Use Disorder and Anxiety: Bridging Psychiatric, Psychological, and Neurobiological Perspectives. Alcohol Res 2019; 40 (01) 4-12
- 59 Gorka SM, Phan KL. Impact of anxiety symptoms and problematic alcohol use on error-related brain activity. Int J Psychophysiol 2017; 118: 32-39
- 60 Pollak CP, Bright D. Caffeine consumption and weekly sleep patterns in US seventh-, eighth-, and ninth-graders. Pediatrics 2003; 111 (01) 42-46
- 61 Ferré S, O'Brien MC. Alcohol and caffeine: The perfect storm. J Caffeine Res 2011; 1 (03) 153-162
- 62 Novati A, Roman V, Cetin T. et al. Chronically restricted sleep leads to depression-like changes in neurotransmitter receptor sensitivity and neuroendocrine stress reactivity in rats. Sleep 2008; 31 (11) 1579-1585
- 63 Boden JM, Fergusson DM. Alcohol and depression. Addiction 2011; 106 (05) 906-914
- 64 Al-Maddah EM, Al-Dabal BK, Khalil MS. Prevalence of sleep deprivation and relation with depressive symptoms among medical residents in King Fahd University Hospital, Saudi Arabia. Sultan Qaboos Univ Med J 2015; 15 (01) e78-e84
- 65 Ghanbari I, Taheri H, Sohrabi M. The Effects of 24-hours sleep Deprivation compared with the acute effects of alcohol and hypoxia. Ann Appl Sport Sci 2019; 1: 27-38
- 66 Karni A, Meyer G, Rey-Hipolito C. et al. The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proc Natl Acad Sci U S A 1998; 95 (03) 861-868
- 67 Elmenhorst D, Elmenhorst EM, Luks N. et al. Performance impairment during four days partial sleep deprivation compared with the acute effects of alcohol and hypoxia. Sleep Med 2009; 10 (02) 189-197
- 68 Vidyasagar J, Karunakar N, Reddy MS, Rajnarayana K, Surender T, Krishna DR. Oxidative stress and antioxidant status in acute organophosphorus insecticide poisoning. Indian J Pharmacol 2004; 37: 76-79
- 69 Akomolafe SF. The effects of caffeine, caffeic acid, and their combination on acetylcholinesterase, adenosine deaminase and arginase activities linked with brain function. J Food Biochem 2017; 41 (05) 12401
- 70 Londzin P, Zamora M, Kąkol B, Taborek A, Folwarczna J. Potential of Caffeine in Alzheimer's Disease-A Review of Experimental Studies. Nutrients 2021; 13 (02) 537
- 71 Collado-Mateo D, Lavín-Pérez AM, Merellano-Navarro E, Coso JD. Effect of Acute Caffeine Intake on the Fat Oxidation Rate during Exercise: A Systematic Review and Meta-Analysis. Nutrients 2020; 12 (12) 3603
- 72 Sethi JK, Hotamisligil GS. Metabolic Messengers: tumour necrosis factor. Nat Metab 2021; 3 (10) 1302-1312
- 73 Roberto M, Patel RR, Bajo M. Cytokines in the CNS. Handb Exp Pharmacol 2018; 248: 397-431
- 74 Garbarino S, Lanteri P, Bragazzi NL, Magnavita N, Scoditti E. Role of sleep deprivation in immune-related disease risk and outcomes. Commun Biol 2021; 4 (01) 1304
- 75 Gano A, Pautassi RM, Doremus-Fitzwater TL, Deak T. Conditioned effects of ethanol on the immune system. Exp Biol Med (Maywood) 2017; 242 (07) 718-730
- 76 Suryanarayanan A, Carter JM, Landin JD, Morrow AL, Werner DF, Spigelman I. Role of interleukin-10 (IL-10) in regulation of GABAergic transmission and acute response to ethanol. Neuropharmacology 2016; 107: 181-188
- 77 Marshall SA, McClain JA, Kelso ML, Hopkins DM, Pauly JR, Nixon K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiol Dis 2013; 54: 239-251
- 78 Felger JC. Imaging the role of inflammation in mood and anxiety-related disorders. Curr Neuropharmacol 2018; 16 (05) 533-558
- 79 Jelic S, Lederer DJ, Adams T. et al. Vascular inflammation in obesity and sleep apnea. Circulation 2010; 121 (08) 1014-1021
Address for correspondence
Publication History
Received: 07 February 2023
Accepted: 17 July 2023
Article published online:
20 February 2024
© 2024. Brazilian Sleep Association. This is an open access article published by Thieme under the terms of the Creative Commons Attribution-NonDerivative-NonCommercial License, permitting copying and reproduction so long as the original work is given appropriate credit. Contents may not be used for commercial purposes, or adapted, remixed, transformed or built upon. (https://creativecommons.org/licenses/by-nc-nd/4.0/)
Thieme Revinter Publicações Ltda.
Rua do Matoso 170, Rio de Janeiro, RJ, CEP 20270-135, Brazil
-
References
- 1 Kamath J, Virdi S, Winokur A. Sleep disturbances in schizophrenia. Psychiatr Clin North Am 2015; 38 (04) 777-792
- 2 Feeley CA, Turner-Henson A, Christian BJ. et al. Sleep quality, stress, caregiver burden, and quality of life in maternal caregivers of young children with bronchopulmonary dysplasia. J Pediatr Nurs 2014; 29 (01) 29-38
- 3 Thakkar MM, Sharma R, Sahota P. Alcohol disrupts sleep homeostasis. Alcohol 2015; 49 (04) 299-310
- 4 Younes M, Hanly PJ. Immediate postarousal sleep dynamics: an important determinant of sleep stability in obstructive sleep apnea. J Appl Physiol 2016; 120 (07) 801-808
- 5 Clark I, Landolt HP. Coffee, caffeine, and sleep: A systematic review of epidemiological studies and randomized controlled trials. Sleep Med Rev 2017; 31: 70-78
- 6 Cappuccio FP, D'Elia L, Strazzullo P, Miller MA. Quantity and quality of sleep and incidence of type 2 diabetes: a systematic review and meta-analysis. Diabetes Care 2010; 33 (02) 414-420
- 7 McLellan TM, Caldwell JA, Lieberman HR. A review of caffeine's effects on cognitive, physical and occupational performance. Neurosci Biobehav Rev 2016; 71: 294-312
- 8 Southward K, Rutherfurd-Markwick KJ, Ali A. The effect of acute caffeine ingestion on endurance performance: a systematic review and meta- analysis. Sports Med 2018; 48 (08) 1913-1928
- 9 Borota D, Murray E, Keceli G. et al. Post-study caffeine administration enhances memory consolidation in humans. Nat Neurosci 2014; 17 (02) 201-203
- 10 Sane RM, Jadhav PR, Subhedar SN. The acute effects of decaffeinated versus caffeinated coffee on reaction time, mood and skeletal muscle strength. J Basic Clin Physiol Pharmacol 2019; 30 (05) 12
- 11 Watson EJ, Coates AM, Kohler M, Banks S. Caffeine consumption and sleep quality in Australian adults. Nutrients 2016; 8 (08) 11-17
- 12 Howell S, Reid JL, Hammond D, McCrory C, Dubin JA, Leatherdale ST. Use of caffeinated energy drinks among secondary school students in Ontario: Prevalence and correlates of using energy drinks and mixing with alcohol. Can J Public Health 2017; 25: 106
- 13 Einöther SJ, Giesbrecht T. Caffeine as an attention enhancer: reviewing existing assumptions. Psychopharmacology (Berl) 2013; 225 (02) 251-274
- 14 Kang S, Yoo H, Chung S, Kim C. Eveningness, sleep patterns, daytime sleepiness and fatigue in Korean male adolescents. Sleep Med Psychophysiol 2012; 19: 89-96
- 15 Boivin DB, Boudreau P. Impacts of shift work on sleep and circadian rhythms. Pathol Biol (Paris) 2014; 62 (05) 292-301
- 16 Meerlo P, Havekes R, Steiger A. Chronically restricted or disrupted sleep as a causal factor in the development of depression. Curr Top Behav Neurosci 2015; 25: 459-481
- 17 Malish S, Arastu F, O'Brien LM. A preliminary study of new parents, sleep disruption, and driving: a population at risk?. Matern Child Health J 2016; 20 (02) 290-297
- 18 Ryu HS, Lee SA, Lee GH, Chung YS, Kim WS. Subjective apnoea symptoms are associated with daytime sleepiness in patients with moderate and severe obstructive sleep apnoea: a retrospective study. Clin Otolaryngol 2016; 41 (04) 395-401
- 19 Welsh T, Mai K, Ellis A, Christian M. Overcoming the effects of sleep deprivation on unethical behavior: an extension of integrated self-control theory. J Exp Soc Psychol 2018; 76: 142-154
- 20 Slama H, Chylinski D, Deliens G, Leproult R, Schmitz R, Peigneux P. Sleep deprivation triggers cognitive control impairments in task-goal switching. Sleep Epub. 2017; 14: 33-39
- 21 Barner C, Altgassen M, Born J, Diekelmann S. Effects of sleep on the realization of complex plans. J Sleep Res 2019; 28 (01) e12655
- 22 Thakkar M, Mallick BN. Effect of REM sleep deprivation on rat brain acetylcholinesterase. Pharmacol Biochem Behav 1991; 39 (01) 211-214
- 23 Paul W, John MH, Brieann CS, Devon AG, Kimberly AH, Hans VD. Sleep Deprivation diminishes attentional control effectiveness and impairs flexible adaptation to changing conditions. Sci Rep 2017; 7 (01) 1-9
- 24 Krause AJ, Simon EB, Mander BA. et al. The sleep-deprived human brain. Nat Rev Neurosci 2017; 18 (07) 404-418
- 25 Zhao SZ, Wang MP, Viswanath K. et al. Short sleep duration and insomnia symptoms were associated with lower happiness levels in Chinese adults. Int J Environ Res Public Health 2019; 16 (12) 2079
- 26 Ali T, Choe J, Awab A, Wagener TL, Orr WC. Sleep, immunity and inflammation in gastrointestinal disorders. World J Gastroenterol 2013; 19 (48) 9231-9239
- 27 Hurtado-Alvarado G, Domínguez-Salazar E, Pavon L, Velázquez-Moctezuma J, Gómez-González B. Blood brain barrier disruption induced by chronic sleep loss: low-grade inflammation may be the link. J Immunol Res 2016; 2016: 4576012
- 28 Wells G, Becker JC, Teng J. et al. Validation of the 28-joint Disease Activity Score (DAS28) and European League Against Rheumatism response criteria based on C-reactive protein against disease progression in patients with rheumatoid arthritis, and comparison with the DAS28 based on erythrocyte sedimentation rate. Ann Rheum Dis 2009; 68 (06) 954-960
- 29 Rawat R, Humphrey JH, Mutasa K, Ntozini R, Stoltzfus RJ. Short communication: predicting adverse HIV-related outcomes in a resource-limited setting: use of the inflammation marker α(1)-acid glycoprotein. AIDS Res Hum Retroviruses 2010; 26 (11) 1171-1174
- 30 Neurath F. Mesenchymal stem cells for chronic wound healing. Nat Rev Immunol 2014; 14: 329
- 31 Ogbutor UG, Nwangwa EK, Nwogueze BC. et al. Proinflammatory and Anti-inflammatory Cytokine Response to Isometric Handgrip Exercise and the Effects of Duration and Intensity of the Isometric Efforts in Prehypertensive Participants. J Chiropr Med 2022; 21 (03) 177-186
- 32 Barnes TC, Anderson ME, Moots RJ. The many faces of interleukin-6: the role of IL-6 in inflammation, vasculopathy, and fibrosis in systemic sclerosis. Int J Rheumatol 2011; 2011: 721608
- 33 Gabay C. Interleukin-6 and chronic inflammation. Arthritis Res Ther 2006; 8 (Suppl. 02) S3
- 34 Javad-Moosavi BZ, Nasehi M, Vaseghi S, Jamaldini SH, Zarrindast MR. Activation and inactivation of nicotinic receptors in the dorsal hippocampal region restored negative effects of total (TSD) and REM sleep deprivation (RSD) on memory acquisition, locomotor activity and pain perception. Neuroscience 2020; 433: 200-211
- 35 Rezaie M, Nasehi M, Vaseghi S. et al. The interaction effect of sleep deprivation and cannabinoid type 1 receptor in the CA1 hippocampal region on passive avoidance memory, depressive-like behavior and locomotor activity in rats. Behav Brain Res 2021; 396: 112901
- 36 Irwin MR, Opp MR, Mark RO. Sleep Health: Reciprocal Regulation of Sleep and Innate Immunity. Neuropsychopharmacology 2017; 42 (01) 129-155
- 37 Ferrucci L, Fabbri E, Sarwan N. Inflammageing: chronic inflammation in ageing, cardiovascular disease, and frailty. Nat Rev Cardiol 2018; 15 (09) 505-522
- 38 Shearer WT, Reuben JM, Mullington JM. et al. Soluble TNF-alpha receptor 1 and IL-6 plasma levels in humans subjected to the sleep deprivation model of spaceflight. J Allergy Clin Immunol 2001; 107 (01) 165-170
- 39 Andreakos E, Bierhaus A, Akerstedt T. Short Communication: Sleep loss activates cellular markers of inflammation. Brain Behav Immun 2017; 24 (01) 54-57
- 40 Fernández-Solà J, Planavila P. Cardiovascular risks and benefits of moderate and heavy alcohol consumption. Nat Rev Cardiol 2015; 12 (10) 576-587
- 41 Bishehsari F, Magno E, Swanson G. et al. Alcohol and Gut-Derived Inflammation. Alcohol Res 2017; 38 (02) 163-171
- 42 Desingu R, Balakrishnan S, Shamsher SK, Bhawna L. Animal models of anxiety: a review. Int J Basic Clin Pharmacol 2023; 12 (01) 134-141
- 43 Habibur R, Muralidharan P, Sivaraman D, Dipankar S. Continuous sleep deprivation for 5 days produces loss of memory in mice and may be a cause of Alzheimer's disease. Ann Biol Res 2010; 1 (04) 185-193
- 44 Silva RH, Kameda SR, Carvalho RC. et al. Anxiogenic effect of sleep deprivation in the elevated plus-maze test in mice. Psychopharmacology (Berl) 2004; 176 (02) 115-122
- 45 Sarter M, Bodewitz G, Stephens DN. Attenuation of scopolamine-induced impairment of spontaneous alteration behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology (Berl) 1988; 94 (04) 491-495
- 46 Komada M, Takao K, Miyakawa T. Elevated plus maze for mice. J Vis Exp 2008; (22) 1088
- 47 Hart PC, Bergner CL, Smolinsky AN. et al. Experimental Models of Anxiety for Drug Discovery and Brain Research. In: Proetzel G, Wiles MV. eds. Mouse Models for Drug Discovery. New York, NY: Springer New York; 2016: 271-291
- 48 Campos AC, Fogaça MV, Aguiar DC, Guimarães FS. Animal models of anxiety disorders and stress. Br J Psychiatry 2013; 35 (Suppl. 02) S101-S111
- 49 Toth I, Neumann ID. Animal models of social avoidance and social fear. Cell Tissue Res 2013; 354 (01) 107-118
- 50 Onaolapo OJ, Onaolapo AY, Akanmu MA, Olayiwola G. Foraging enrichment modulates open field response to monosodium glutamate in mice. Ann Neurosci 2015; 22 (03) 162-170
- 51 Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 1979; 95 (02) 351-358
- 52 Ellman GL, Courtney KD, Andres Jr V, Feather-Stone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 1961; 7: 88-95
- 53 Peng Z, Dai C, Ba Y, Zhang L, Shao Y, Tian J. Effect of Sleep Deprivation on the Working Memory-Related N2-P3 Components of the Event-Related Potential Waveform. Front Neurosci 2020; 14: 469
- 54 Vedder LC, Hall JM, Jabrouin KR, Savage LM. Interactions between chronic ethanol consumption and thiamine deficiency on neural plasticity, spatial memory, and cognitive flexibility. Alcohol Clin Exp Res 2015; 39 (11) 2143-2153
- 55 Chen X, Zhang L, Yang D. et al. Effects of Caffeine on Event-Related Potentials and Neuropsychological Indices After Sleep Deprivation. Front Behav Neurosci 2020; 14: 108
- 56 Chang YS, Chen HL, Wu YH, Hsu CY, Liu CK, Hsu C. Rotating night shifts too quickly may cause anxiety and decreased attentional performance, and impact prolactin levels during the subsequent day: a case control study. BMC Psychiatry 2014; 14 (01) 218
- 57 Holmes A, Fitzgerald PJ, MacPherson KP. et al. Chronic alcohol remodels prefrontal neurons and disrupts NMDAR-mediated fear extinction encoding. Nat Neurosci 2012; 15 (10) 1359-1361
- 58 Anker JJ, Kushner MG. Co-Occurring Alcohol Use Disorder and Anxiety: Bridging Psychiatric, Psychological, and Neurobiological Perspectives. Alcohol Res 2019; 40 (01) 4-12
- 59 Gorka SM, Phan KL. Impact of anxiety symptoms and problematic alcohol use on error-related brain activity. Int J Psychophysiol 2017; 118: 32-39
- 60 Pollak CP, Bright D. Caffeine consumption and weekly sleep patterns in US seventh-, eighth-, and ninth-graders. Pediatrics 2003; 111 (01) 42-46
- 61 Ferré S, O'Brien MC. Alcohol and caffeine: The perfect storm. J Caffeine Res 2011; 1 (03) 153-162
- 62 Novati A, Roman V, Cetin T. et al. Chronically restricted sleep leads to depression-like changes in neurotransmitter receptor sensitivity and neuroendocrine stress reactivity in rats. Sleep 2008; 31 (11) 1579-1585
- 63 Boden JM, Fergusson DM. Alcohol and depression. Addiction 2011; 106 (05) 906-914
- 64 Al-Maddah EM, Al-Dabal BK, Khalil MS. Prevalence of sleep deprivation and relation with depressive symptoms among medical residents in King Fahd University Hospital, Saudi Arabia. Sultan Qaboos Univ Med J 2015; 15 (01) e78-e84
- 65 Ghanbari I, Taheri H, Sohrabi M. The Effects of 24-hours sleep Deprivation compared with the acute effects of alcohol and hypoxia. Ann Appl Sport Sci 2019; 1: 27-38
- 66 Karni A, Meyer G, Rey-Hipolito C. et al. The acquisition of skilled motor performance: fast and slow experience-driven changes in primary motor cortex. Proc Natl Acad Sci U S A 1998; 95 (03) 861-868
- 67 Elmenhorst D, Elmenhorst EM, Luks N. et al. Performance impairment during four days partial sleep deprivation compared with the acute effects of alcohol and hypoxia. Sleep Med 2009; 10 (02) 189-197
- 68 Vidyasagar J, Karunakar N, Reddy MS, Rajnarayana K, Surender T, Krishna DR. Oxidative stress and antioxidant status in acute organophosphorus insecticide poisoning. Indian J Pharmacol 2004; 37: 76-79
- 69 Akomolafe SF. The effects of caffeine, caffeic acid, and their combination on acetylcholinesterase, adenosine deaminase and arginase activities linked with brain function. J Food Biochem 2017; 41 (05) 12401
- 70 Londzin P, Zamora M, Kąkol B, Taborek A, Folwarczna J. Potential of Caffeine in Alzheimer's Disease-A Review of Experimental Studies. Nutrients 2021; 13 (02) 537
- 71 Collado-Mateo D, Lavín-Pérez AM, Merellano-Navarro E, Coso JD. Effect of Acute Caffeine Intake on the Fat Oxidation Rate during Exercise: A Systematic Review and Meta-Analysis. Nutrients 2020; 12 (12) 3603
- 72 Sethi JK, Hotamisligil GS. Metabolic Messengers: tumour necrosis factor. Nat Metab 2021; 3 (10) 1302-1312
- 73 Roberto M, Patel RR, Bajo M. Cytokines in the CNS. Handb Exp Pharmacol 2018; 248: 397-431
- 74 Garbarino S, Lanteri P, Bragazzi NL, Magnavita N, Scoditti E. Role of sleep deprivation in immune-related disease risk and outcomes. Commun Biol 2021; 4 (01) 1304
- 75 Gano A, Pautassi RM, Doremus-Fitzwater TL, Deak T. Conditioned effects of ethanol on the immune system. Exp Biol Med (Maywood) 2017; 242 (07) 718-730
- 76 Suryanarayanan A, Carter JM, Landin JD, Morrow AL, Werner DF, Spigelman I. Role of interleukin-10 (IL-10) in regulation of GABAergic transmission and acute response to ethanol. Neuropharmacology 2016; 107: 181-188
- 77 Marshall SA, McClain JA, Kelso ML, Hopkins DM, Pauly JR, Nixon K. Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: The importance of microglia phenotype. Neurobiol Dis 2013; 54: 239-251
- 78 Felger JC. Imaging the role of inflammation in mood and anxiety-related disorders. Curr Neuropharmacol 2018; 16 (05) 533-558
- 79 Jelic S, Lederer DJ, Adams T. et al. Vascular inflammation in obesity and sleep apnea. Circulation 2010; 121 (08) 1014-1021