
RSS-Feed abonnieren
DOI: 10.1055/s-0042-1742774
The Relationship between Contralateral Suppression of Transient Evoked Otoacoustic Emission and Unmasking of Speech Evoked Auditory Brainstem Response


Abstract
Introduction Several studies have shown that efferent pathways of the auditory system improve perception of speech-in-noise. But, the majority of investigations assessing the role of efferent pathways on speech perception have used contralateral suppression of otoacoustic emissions as a measure of efferent activity. By studying the effect of efferent activity on the speech-evoked auditory brainstem response (ABR), some more light could be shed on the effect of efferent pathways on the encoding of speech in the auditory pathway.
Objectives To investigate the relationship between contralateral suppression of transient evoked otoacoustic emission (CSTEOAE) and unmasking of speech ABR.
Methods A total of 23 young adults participated in the study. The CSTEOAE was measured using linear clicks at 60 dB peSPL and white noise at 60 dB sound pressure level (SPL). The speech ABR was recorded using the syllable /da/ at 80 dB SPL in quiet, ipsilateral noise, and binaural noise conditions. In the ipsilateral noise condition, white noise was presented to the test ear at 60 dB SPL, and, in the binaural noise condition, two separate white noises were presented to both ears.
Results The F0 amplitude of speech ABR was higher in quiet condition; however, the mean amplitude of F0 was not significantly different across conditions. Correlation analysis showed a significant positive correlation between the CSTEOAE and the magnitude of unmasking of F0 amplitude of speech ABR.
Conclusions The findings of the present study suggests that the efferent pathways are involved in speech-in-noise processing.
Introduction
The human auditory system consists of afferent and efferent pathways that interact with each other while processing the auditory information.[1] The efferent pathways of the auditory system are believed to aid in selective attention,[2] [3] protect the inner ear from auditory fatigability and acoustic trauma,[4] [5] [6] and improve the coding of signals embedded in noise,[7] [8] [9] speech perception in noise,[10] [11] [12] [13] and sound localization in noise.[14] The role of efferent auditory pathways on speech perception in noise has received greater attention among researchers,[10] [11] [12] [13] [15] [16] [17] [18] [19] [20] [21] [22] but their functional role in speech perception is not understood.
Several studies have investigated the role of efferent auditory pathways on the perception of speech in noise. Earlier, investigations were performed among individuals who had undergone transection of the olivocochlear bundle during vestibular neurectomy.[11] [17] These investigations elicited efferent activity of the auditory system by presenting noise to the contralateral ear of participants (referred to as contralateral noise). Findings of the above investigations showed a significant improvement in speech identification score (5–10%) among normally hearing individuals when noise was presented to the contralateral ear. In contrast, no such improvement in speech identification score was observed among individuals who had undergone vestibular neurectomy. These findings revealed the significance of efferent pathways for the perception of speech in noise. Thus, the efferent pathways are assumed to improve speech perception in noise.
On the other hand, most studies have investigated the role of efferent pathways non-invasively by assessing the relationship between contralateral suppression of otoacoustic emissions (CSOAEs) and perception of speech in noise. Several investigations have reported a significant correlation between speech perception in noise and the magnitude of CSOAE.[10] [11] [12] [13] [18] [19] In contrast, studies have also reported no relationship between speech perception in noise and the CSOAEs.[20] [21] [22] [23] [24] The discrepancy in findings across investigations could be due to differences in tasks used to measure the CSOAE and speech perception in noise. Though no conclusive evidence is available, the efferent pathways are believed to improve the encoding of speech in the presence of noise. However, the CSOAE, which is commonly used for the assessment of the efferent system, measures the magnitude of efferent activity and does not provide information about the encoding of speech in the auditory system.
Speech evoked auditory brainstem response (speech ABR) is a useful tool to investigate the encoding of speech in the auditory system, and it provides reliable information about the neural coding of speech sounds.[25] It has been widely used to investigate the encoding of speech among elderly adults and children with auditory processing deficits as well as to explain their speech perception difficulties.[26] [27] By studying the effect of contralateral noise on the speech ABR, we could understand the role of efferent pathways on the encoding of speech in the presence of noise at the neural level. Recently, an investigation showed no significant effect of contralateral noise on speech ABR in quiet and in noise. In addition, no significant correlation was found between CSOAE and unmasking of speech ABR.[28] The findings of the above-mentioned study suggest that efferent activity may not have any effect on the neural encoding of speech in the presence of noise. However, other similar studies are required before generalizing the findings of the above-mentioned investigation. The present study was aimed to investigate the relationship between contralateral suppression of speech ABR and contralateral suppression of transient evoked otoacoustic emission (CSTEOAE).
Material and Methods
Participants
A total of 23 adults aged between 18 and 40 years old (mean age: 29 years) participated in the study. All the participants had hearing sensitivity within normal limits in both ears. The immittance evaluation showed ‘A’ type tympanogram with ipsilateral and contralateral acoustic reflex present for pure tones at normal levels. In addition, the contralateral acoustic reflex threshold for white noise was greater than 70 dB SPL. None of the participants had difficulty understanding speech in quiet or noise. In addition, none of the participants had a history of otological or neurological dysfunction, metabolic disorders (diabetes and hypertension), or exposure to hazardous noise or ototoxic drugs. Finally, all the participants had TEOAEs present in both ears for non-linear clicks at 80 dB SPL. The institutional ethics committee approved the study, and an informed consent form was obtained from all the participants before joining this study.
Stimuli
The consonant-vowel syllable /da/, spoken by a female native speaker of Kannada, was used to elicit speech ABR. The waveform and spectrum of stimulus used in the present study are shown in [Fig. 1]. It included a stop burst in the beginning, followed by a harmonically rich and spectrally dynamic formant transition. The stimulus duration was 232.5 milliseconds, the fundamental frequency was 162 Hz, and the first formant frequency (F1) was 820 Hz. White noise was used to elicit the efferent activity, and it was generated using the Praat software (Institute of Phonetic Sciences, University of Amsterdam, The Netherlands).[29]

Procedure
Recording of TEOAEs
During the recording of TEOAEs, participants were made to sit comfortably on a reclining chair. The Echoport ILO 292 OAE analyser (Otodynamics Ltd., Hatfield, UK) and the ILO V6 computer software (Otodynamics Ltd., Hatfield, UK) were used for the recording of TEOAEs. Initially, the TEOAE was recorded using non-linear clicks at 80 dB peak sound pressure level (SPL) to confirm the presence of TEOAE. The TEOAEs were considered as present when the global signal-to-noise ratio (SNR) and reproducibility were greater than 6 dB SNR and 80%, respectively. Following this, the TEOAEs were recorded using ‘linear clicks’ at 60 dB p.e SPL to measure the contralateral suppression of TEOAE. First, a ‘baseline’ TEOAE was recorded without presenting noise to the contralateral ear of participants. Following this, a second TEOAE was recorded by presenting white noise to the contralateral ear of participants. Finally, an additional baseline TEOAE was obtained at the end of the recording of TEOAEs. The white noise was delivered to the contralateral ear of participants using ER-5A insert phones (Etymotic Research, Elk Grove Village, IL) at 60 dB SPL. The TEOAEs for linear clicks were considered as present when the SNR was greater than 3 dB SNR. The global amplitude of TEOAE and noise floor were obtained from the ILO V6 software. The magnitude of CSTEOAE was computed by subtracting the amplitude of TEOAE with contralateral noise from the amplitude of baseline TEOAE.
Recording of Speech Evoked ABR
The speech ABR was recorded using the IHS Smart EP evoked potential system version 3.92 (Intelligent Hearing Systems, Miami, FL, USA). During the recording of speech ABR, the participants were made to sit comfortably on a reclining chair. They were instructed to relax and minimize extraneous body movements to reduce unwanted artifacts. The electroencephalogram (EEG) was differentially recorded from the scalp using gold-plated disc electrodes. The non-inverting electrode was placed on the vertex (Cz), inverting electrode on the test ear mastoid (A2), and the ground electrode was placed on the mastoid of the non-test ear (A1). To elicit the speech ABR, stimuli of single polarity were presented to the right ear of participants using Etymotic ER-3A (Etymotic Research, Elk Grove Village, IL) insert earphones. A total of 2,000 artifact-free responses were collected and averaged to obtain the averaged waveform in each recording. Initially, the speech ABR was recorded in quiet condition. Following this, the speech ABR was recorded in noise conditions by presenting white noise to the test ear and then both the ears, and the order of noise conditions was randomized. The white noise was delivered to the contralateral ear of participants using ER-5A insert phones at 60 dB SPL. A short break of 5 to 10 minutes was provided between the recordings as required by the participants. Two recordings were obtained in each condition, once for rarefaction and condensation polarities. The recording parameters used in the present study for the recording of speech ABR are shown in [Table 1].
Using the speech ABR waveforms for rarefaction and condensation polarities, the sum and the difference waveforms were obtained. Adding and subtracting the waveforms of rarefaction and condensation polarities selectively enhances the amplitude of components of speech ABR.[30] The sum and difference waveform were subjected to fast Fourier transform (FFT) analysis to measure the amplitude at the F0 and F1 components of speech ABR respectively. For this purpose, a 50-millisecond segment of the averaged waveform was extracted in the post-stimulus response waveform (88–138 milliseconds). To increase frequency resolution in the FFT spectrum, the length of the extracted waveform was increased to 1,024 points by zero-padding. From the FFT spectrum, the peak amplitude at frequencies between 150 and 174 Hz and 740 and 900 Hz was noted, referred to as peak F0 and peak F1 amplitudes, respectively. In addition, the average F0 and F1 amplitude were computed by averaging the amplitude at frequencies between 150 and 174 Hz and 740 and 900 Hz, respectively. The difference in amplitude of F0 between ipsilateral noise and binaural noise conditions was considered contralateral suppression (unmasking) of speech ABR.
Statistical Analysis
Initially, the amplitude of TEOAE in baseline and contralateral noise conditions, the magnitude of CSTEOAE, the amplitude of F0 and F1 of speech ABR across conditions, and the magnitude of unmasking of the amplitude of F0 were subjected to the Shapiro-Wilk test to check for normal distribution. The paired samples ‘t’ test was performed to investigate the effect of conditions on the amplitude of TEOAE. A repeated measure analysis of variance (ANOVA) was performed to investigate the effect of conditions (quiet, ipsilateral noise, and binaural noise) on the F0 amplitude of speech ABR. A Pearson correlation analysis was performed to investigate the relationship between CSTEOAE and the unmasking of the amplitude of speech ABR.
Results
The TEOAE data of 19 participants were available for statistical analysis; in the remaining four participants, the TEOAE was judged to be absent due to high noise floor. The mean amplitude of TEOAE in the baseline condition was 8.2 dB SPL (standard deviation [SD = 6.3]), and, in the contralateral noise condition, it was 6.9 dB SPL (SD = 6.5). The mean contralateral suppression of TEOAE was 1.3 dB (SD = 0.8). The amplitude of TEOAE in the baseline condition was higher than the amplitude of TEOAE in the contralateral noise condition. The Shapiro-Wilk test showed that the TEOAE amplitude difference between baseline and contralateral noise conditions was normally distributed (W = 0.97, p = 0.768). To investigate if the mean amplitudes are significantly different between conditions (baseline and contralateral noise), a paired samples ‘t’ test was performed. It showed a significant effect of conditions on the amplitude of TEOAEs (t [13] = 6.184, p < 0.001). Thus, the reduction in the amplitude of TEOAE in contralateral noise condition was significant.
The speech ABR was present in 15 individuals in quiet and ipsilateral noise conditions and in 14 individuals in binaural noise condition. In the remaining eight participants, the speech ABR was found to be absent. The amplitude of F0 was above the noise floor for all participants across conditions. In contrast, the amplitude of F1 in quiet and noise conditions was above the noise floor in 7 and 8 participants, respectively. Since the amplitude of F1 was measurable only in 50% of the participants, it was not considered for further statistical analysis. [Fig. 2] shows grand averaged waveforms of speech ABR in quiet and noise conditions. [Fig. 3] shows the spectrum of pre-stimulus activity and response waveform in quiet and noise conditions. [Table 2] shows the mean peak F0 amplitude of the and average F0 amplitude for transition and sustained portion of speech ABR across quiet and noise conditions. The mean peak F0 and average F0 amplitude were highest in quiet condition and lowest in binaural noise condition. In noise, the mean amplitude of F0 was similar in both ipsilateral noise and binaural noise conditions. Furthermore, among 8 (57.1%) participants, the amplitude of F0 in binaural noise condition was greater than in ipsilateral noise condition.


The Shapiro-Wilk test showed that the peak F0 amplitude of the transition portion and the peak F0 amplitude and average F0 of the sustained portion of speech ABR across conditions was normally distributed (p > 0.05). But the average F0 amplitude of the transition portion in quiet condition was not normally distributed (W = 0.819, p = 0.009). To investigate if the F0 amplitudes are significantly different across conditions, a repeated-measures ANOVA was performed separately for the peak F0 amplitude of the transition and sustained portions and average F0 amplitude of the sustained portion of speech ABR with conditions (quiet, ipsilateral noise, and binaural noise) as repeated measures. It showed no significant effect of conditions on the peak F0 amplitude of the transition portion (F [2.26] = 0.906, p = 0.417), peak F0 amplitude (F [2.26] = 0.42, p = 0.662), and average F0 amplitude (F [1.434,18.644] = 0.44, p = 0.586) of the sustained portion. The Friedman test was performed for the average F0 amplitude of the transition portion of speech ABR with conditions (quiet, ipsilateral noise, and binaural noise) as repeated measures. It showed no significant effect of conditions on the average F0 amplitude of the transition portion (χ2[2.26] = 0.571, p = 0.751).
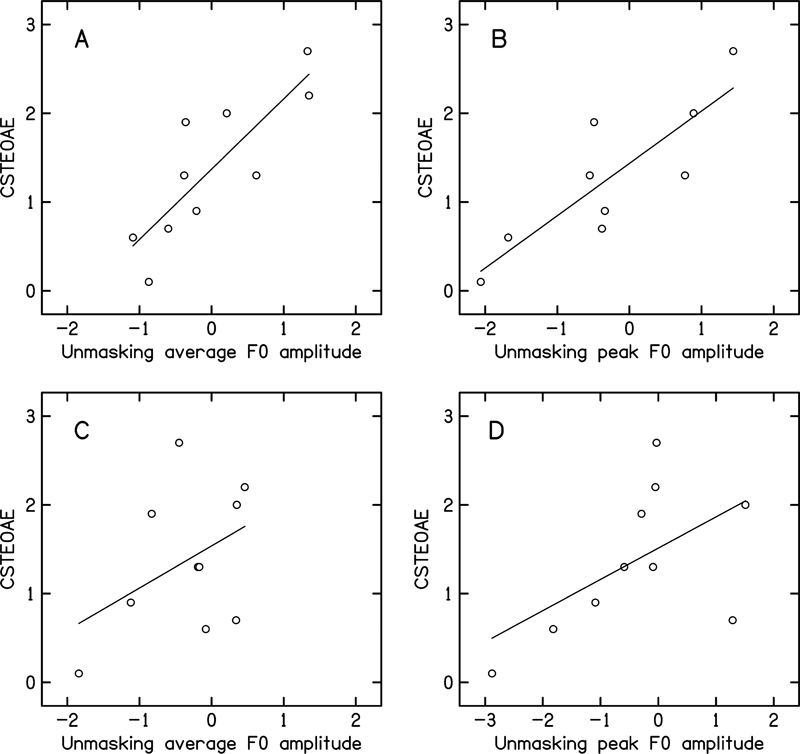
The data was subjected to correlation analysis to investigate the relationship between CSTEOAE and unmasking of the amplitude of F0 of speech ABR. The Shapiro-Wilk test showed that the magnitude of unmasking of the amplitude of peak F0 and average F0 of speech ABR was normally distributed for the sustained portion (peak F0 [W = 0.9053, p = 0.158]; average F0 [W = 0.9388, p = 0.403]), but was not normally distributed for the transition portion (peak F0 [W = 0.8231, p = 0.01]; average F0 [W = 0.7049, p < 0.001]). The Pearson correlation analysis was performed to investigate the relationship between CSTEOAE and unmasking of the F0 amplitude of speech ABR for the sustained portion. The Spearman correlation analysis was performed to investigate the relationship between CSTEOAE and unmasking of the F0 amplitude of speech ABR for the transition portion. The results of the correlation analysis are shown in [Table 3]. The results showed a significant positive correlation between the magnitude of CSTEOAE and the magnitude of unmasking of the peak F0 amplitude (both sustained and transition portion) and the average F0 amplitude of speech ABR of sustained portion. [Fig. 4] shows the scatter plot with trend line showing the relationship between CSTEOAE and the magnitude of unmasking of the amplitude of ABR.
Abbreviation: TEOAE, transient evoked otoacoustic emission.
Note: * p < 0.05, ** p < 0.01, *** p < 0.001.
Discussion
The findings of the present study showed a significant reduction in the amplitude of TEOAE when white noise was presented to the contralateral ear compared with the baseline condition. These results are consistent with the findings of several investigations.[12] [18] [20] [31] Further, the mean CSTEOAE obtained in the present study was comparable to the results of earlier investigations.[31] On the other hand, the mean F0 amplitude of speech ABR was highest in quiet condition than in noise condition. Several investigations have reported a similar finding.[28] [32] [33] The reduction of the F0 amplitude in noise conditions could be attributed to the masking effects of noise on the speech ABR. Further, the mean amplitude of F0 was similar in both binaural and ipsilateral noise conditions. These findings are comparable to the findings of earlier investigation.[28] In addition, the amplitude of F0 was higher in binaural noise condition compared with ipsilateral noise condition among 57% of the participants. The higher amplitude of F0 in binaural noise compared with ipsilateral noise condition reflects the unmasking effects of efferent activity in the auditory system.
The present study showed a significant positive correlation between contralateral suppression of the amplitude of F0 speech ABR and TEOAE; that is, participants with stronger efferent activity showed greater unmasking of speech ABR. This finding of the present study contrasts with the results of an earlier investigation,[28] which showed no relationship between the two measures. The positive relationship found in the present study suggested that the efferent pathways are involved in speech-in-noise processing. This finding of the present study is consistent with several investigations that have demonstrated a relationship between the magnitude of efferent activity and speech recognition in noise.[11] [12] [13] [18] [19] However, further investigations are required before generalizing the findings and also to evaluate the reproducibility of the findings of the present study. The limitation of the present study is that perception of speech-in-noise was not measured in the presence and absence of efferent activity. Assessing the relationship between the difference in speech perception in noise scores (between presence and absence of efferent activity) and unmasking of speech ABR could reveal the involvement of efferent pathways in speech-in-noise processing. Furthermore, in the present study, the TEOAEs obtained in linear mode were considered to be present when the SNR was greater than 3 dB. This could be a limitation, as few studies have recommended very high SNR for the detection of small changes in OAE.[34] [35] Finally, another limitation of the present study was the small sample size. Although a total of 23 adults participated in the study, the speech ABR was present only in 14 participants, which could be attributed to individual variability of speech ABR.
Conclusion
The findings of the present study showed that the efferent pathways are involved in speech-in-noise processing. However, further research is required before generalizing the findings of the study.
Conflict of Interests
The authors have no conflict of interests to declare.
-
References
- 1 Lopez-Poveda EA. Olivocochlear efferents in animals and humans: From anatomy to clinical relevance. Front Neurol 2018; 9: 197
- 2 Scharf B, Magnan J, Chays A. On the role of the olivocochlear bundle in hearing: 16 case studies. Hear Res 1997; 103 (1-2): 101-122
- 3 Bell A, Jedrzejczak WW. Muscles in and around the ear as the source of “physiological noise” during auditory selective attention: A review and novel synthesis. Eur J Neurosci 2021; 53 (08) 2726-2739
- 4 Cody AR, Johnstone BM. Acoustically evoked activity of single efferent neurons in the guinea pig cochlea. J Acoust Soc Am 1982; 72 (01) 280-282
- 5 Zheng XY, Henderson D, McFadden SL, Hu BH. The role of the cochlear efferent system in acquired resistance to noise-induced hearing loss. Hear Res 1997; 104 (1-2): 191-203
- 6 Rajan R. Centrifugal pathways protect hearing sensitivity at the cochlea in noisy environments that exacerbate the damage induced by loud sound. J Neurosci 2000; 20 (17) 6684-6693
- 7 Liberman MC. Physiology of cochlear efferent and afferent neurons: direct comparisons in the same animal. Hear Res 1988; 34 (02) 179-191
- 8 May BJ, McQuone SJ. Effects of bilateral olivocochlear lesions on pure-tone intensity discrimination in cats. Aud Neurosci 1995; 1 (04) 385-400
- 9 Otsuka S, Tsuzaki M, Sonoda J, Tanaka S, Furukawa S. A role of medial olivocochlear reflex as a protection mechanism from noise-induced hearing loss revealed in short-practicing violinists. Malmierca MS, editor. PLoS One. 2016; 11 (01) e0146751
- 10 Mertes IB, Johnson KM, Dinger ZA. Olivocochlear efferent contributions to speech-in-noise recognition across signal-to-noise ratios. J Acoust Soc Am 2019; 145 (03) 1529-1540
- 11 Giraud AL, Garnier S, Micheyl C, Lina G, Chays A, Chéry-Croze S. Auditory efferents involved in speech-in-noise intelligibility. Neuroreport 1997; 8 (07) 1779-1783
- 12 Garinis AC, Glattke T, Cone BK. The MOC reflex during active listening to speech. J Speech Lang Hear Res 2011; 54 (05) 1464-1476
- 13 Mishra SK, Lutman ME. Top-down influences of the medial olivocochlear efferent system in speech perception in noise. PLoS One 2014; 9 (01) e85756
- 14 Andéol G, Guillaume A, Micheyl C, Savel S, Pellieux L, Moulin A. Auditory efferents facilitate sound localization in noise in humans. J Neurosci 2011; 31 (18) 6759-6763
- 15 de Boer J, Thornton ARD, Krumbholz K. What is the role of the medial olivocochlear system in speech-in-noise processing?. J Neurophysiol 2012; 107 (05) 1301-1312
- 16 Shastri U, Mythri HM, Kumar UA. Descending auditory pathway and identification of phonetic contrast by native listeners. J Acoust Soc Am 2014; 135 (02) 896-905
- 17 Zeng F-G, Martino KM, Linthicum FH, Soli SD. Auditory perception in vestibular neurectomy subjects. Hear Res 2000; 142 (1-2): 102-112
- 18 Kumar UA, Vanaja CS. Functioning of olivocochlear bundle and speech perception in noise. Ear Hear 2004; 25 (02) 142-146
- 19 Kim S, Frisina RD, Frisina DR. Effects of age on speech understanding in normal hearing listeners: Relationship between the auditory efferent system and speech intelligibility in noise. Speech communication 2006; 48 (07) 855-862
- 20 Narne VK, Kalaiah MK. Involvement of the efferent auditory system for improvement in speech perception in noise. Int J Speech Lang Pathol Audiol 2018; 6: 1-7
- 21 Wagner W, Frey K, Heppelmann G, Plontke SK, Zenner H-P. Speech-in-noise intelligibility does not correlate with efferent olivocochlear reflex in humans with normal hearing. Acta Otolaryngol 2008; 128 (01) 53-60
- 22 Mukari SZ-MS, Mamat WHW. Medial olivocochlear functioning and speech perception in noise in older adults. Audiol Neurotol 2008; 13 (05) 328-334
- 23
Boothalingam S,
Allan C,
Allen P,
Purcell DW.
The medial olivocochlear reflex is unlikely to play a role in listening difficulties
in children. Trends Hear 2019; 23: 2331216519870942
MissingFormLabel
- 24 Stuart A, Butler AK. Contralateral suppression of transient otoacoustic emissions and sentence recognition in noise in young adults. J Am Acad Audiol 2012; 23 (09) 686-696
- 25 Clinard CG, Tremblay KL. Aging degrades the neural encoding of simple and complex sounds in the human brainstem. J Am Acad Audiol 2013; 24 (07) 590-599 , quiz 643–644
- 26 Anderson S, Skoe E, Chandrasekaran B, Zecker S, Kraus N. Brainstem correlates of speech-in-noise perception in children. Hear Res 2010; 270 (1-2): 151-157
- 27 Anderson S, Parbery-Clark A, White-Schwoch T, Kraus N. Auditory brainstem response to complex sounds predicts self-reported speech-in-noise performance. J Speech Lang Hear Res 2013; 56 (01) 31-43
- 28 Smith SB, Cone B. Efferent unmasking of speech-in-noise encoding?. Int J Audiol 2021; 1-10
- 29 Boersma P, Weenink D. Praat: doing phonetics by computer [Computer program]. 2017
- 30 Kumar K, Bhat JS, D'Costa PE, Srivastava M, Kalaiah MK. Effect of stimulus polarity on speech evoked auditory brainstem response. Audiology Res 2014; 3 (01) e8
- 31 Kalaiah MK, Nanchirakal JF, Kharmawphlang L, Noronah SC. Contralateral suppression of transient evoked otoacoustic emissions for various noise signals. Hearing, Balance and Communication 2017; 15 (02) 84-90
- 32 Russo N, Nicol T, Musacchia G, Kraus N. Brainstem responses to speech syllables. Clin Neurophysiol 2004; 115 (09) 2021-2030
- 33 Song JH, Skoe E, Banai K, Kraus N. Perception of speech in noise: neural correlates. J Cogn Neurosci 2011; 23 (09) 2268-2279
- 34 Goodman SS, Mertes IB, Lewis JD, Weissbeck DK. Medial olivocochlear-induced transient-evoked otoacoustic emission amplitude shifts in individual subjects. J Assoc Res Otolaryngol 2013; 14 (06) 829-842
- 35 Jedrzejczak WW, Pilka E, Kochanek K, Skarzynski H. Does the presence of spontaneous components affect the reliability of contralateral suppression of evoked otoacoustic emissions?. Ear Hear 2021; 42 (04) 990-1005
Address for correspondence
Publikationsverlauf
Eingereicht: 22. Juli 2021
Angenommen: 21. Dezember 2021
Artikel online veröffentlicht:
20. April 2022
© 2022. Fundação Otorrinolaringologia. This is an open access article published by Thieme under the terms of the Creative Commons Attribution-NonDerivative-NonCommercial License, permitting copying and reproduction so long as the original work is given appropriate credit. Contents may not be used for commecial purposes, or adapted, remixed, transformed or built upon. (https://creativecommons.org/licenses/by-nc-nd/4.0/)
Thieme Revinter Publicações Ltda.
Rua do Matoso 170, Rio de Janeiro, RJ, CEP 20270-135, Brazil
-
References
- 1 Lopez-Poveda EA. Olivocochlear efferents in animals and humans: From anatomy to clinical relevance. Front Neurol 2018; 9: 197
- 2 Scharf B, Magnan J, Chays A. On the role of the olivocochlear bundle in hearing: 16 case studies. Hear Res 1997; 103 (1-2): 101-122
- 3 Bell A, Jedrzejczak WW. Muscles in and around the ear as the source of “physiological noise” during auditory selective attention: A review and novel synthesis. Eur J Neurosci 2021; 53 (08) 2726-2739
- 4 Cody AR, Johnstone BM. Acoustically evoked activity of single efferent neurons in the guinea pig cochlea. J Acoust Soc Am 1982; 72 (01) 280-282
- 5 Zheng XY, Henderson D, McFadden SL, Hu BH. The role of the cochlear efferent system in acquired resistance to noise-induced hearing loss. Hear Res 1997; 104 (1-2): 191-203
- 6 Rajan R. Centrifugal pathways protect hearing sensitivity at the cochlea in noisy environments that exacerbate the damage induced by loud sound. J Neurosci 2000; 20 (17) 6684-6693
- 7 Liberman MC. Physiology of cochlear efferent and afferent neurons: direct comparisons in the same animal. Hear Res 1988; 34 (02) 179-191
- 8 May BJ, McQuone SJ. Effects of bilateral olivocochlear lesions on pure-tone intensity discrimination in cats. Aud Neurosci 1995; 1 (04) 385-400
- 9 Otsuka S, Tsuzaki M, Sonoda J, Tanaka S, Furukawa S. A role of medial olivocochlear reflex as a protection mechanism from noise-induced hearing loss revealed in short-practicing violinists. Malmierca MS, editor. PLoS One. 2016; 11 (01) e0146751
- 10 Mertes IB, Johnson KM, Dinger ZA. Olivocochlear efferent contributions to speech-in-noise recognition across signal-to-noise ratios. J Acoust Soc Am 2019; 145 (03) 1529-1540
- 11 Giraud AL, Garnier S, Micheyl C, Lina G, Chays A, Chéry-Croze S. Auditory efferents involved in speech-in-noise intelligibility. Neuroreport 1997; 8 (07) 1779-1783
- 12 Garinis AC, Glattke T, Cone BK. The MOC reflex during active listening to speech. J Speech Lang Hear Res 2011; 54 (05) 1464-1476
- 13 Mishra SK, Lutman ME. Top-down influences of the medial olivocochlear efferent system in speech perception in noise. PLoS One 2014; 9 (01) e85756
- 14 Andéol G, Guillaume A, Micheyl C, Savel S, Pellieux L, Moulin A. Auditory efferents facilitate sound localization in noise in humans. J Neurosci 2011; 31 (18) 6759-6763
- 15 de Boer J, Thornton ARD, Krumbholz K. What is the role of the medial olivocochlear system in speech-in-noise processing?. J Neurophysiol 2012; 107 (05) 1301-1312
- 16 Shastri U, Mythri HM, Kumar UA. Descending auditory pathway and identification of phonetic contrast by native listeners. J Acoust Soc Am 2014; 135 (02) 896-905
- 17 Zeng F-G, Martino KM, Linthicum FH, Soli SD. Auditory perception in vestibular neurectomy subjects. Hear Res 2000; 142 (1-2): 102-112
- 18 Kumar UA, Vanaja CS. Functioning of olivocochlear bundle and speech perception in noise. Ear Hear 2004; 25 (02) 142-146
- 19 Kim S, Frisina RD, Frisina DR. Effects of age on speech understanding in normal hearing listeners: Relationship between the auditory efferent system and speech intelligibility in noise. Speech communication 2006; 48 (07) 855-862
- 20 Narne VK, Kalaiah MK. Involvement of the efferent auditory system for improvement in speech perception in noise. Int J Speech Lang Pathol Audiol 2018; 6: 1-7
- 21 Wagner W, Frey K, Heppelmann G, Plontke SK, Zenner H-P. Speech-in-noise intelligibility does not correlate with efferent olivocochlear reflex in humans with normal hearing. Acta Otolaryngol 2008; 128 (01) 53-60
- 22 Mukari SZ-MS, Mamat WHW. Medial olivocochlear functioning and speech perception in noise in older adults. Audiol Neurotol 2008; 13 (05) 328-334
- 23
Boothalingam S,
Allan C,
Allen P,
Purcell DW.
The medial olivocochlear reflex is unlikely to play a role in listening difficulties
in children. Trends Hear 2019; 23: 2331216519870942
MissingFormLabel
- 24 Stuart A, Butler AK. Contralateral suppression of transient otoacoustic emissions and sentence recognition in noise in young adults. J Am Acad Audiol 2012; 23 (09) 686-696
- 25 Clinard CG, Tremblay KL. Aging degrades the neural encoding of simple and complex sounds in the human brainstem. J Am Acad Audiol 2013; 24 (07) 590-599 , quiz 643–644
- 26 Anderson S, Skoe E, Chandrasekaran B, Zecker S, Kraus N. Brainstem correlates of speech-in-noise perception in children. Hear Res 2010; 270 (1-2): 151-157
- 27 Anderson S, Parbery-Clark A, White-Schwoch T, Kraus N. Auditory brainstem response to complex sounds predicts self-reported speech-in-noise performance. J Speech Lang Hear Res 2013; 56 (01) 31-43
- 28 Smith SB, Cone B. Efferent unmasking of speech-in-noise encoding?. Int J Audiol 2021; 1-10
- 29 Boersma P, Weenink D. Praat: doing phonetics by computer [Computer program]. 2017
- 30 Kumar K, Bhat JS, D'Costa PE, Srivastava M, Kalaiah MK. Effect of stimulus polarity on speech evoked auditory brainstem response. Audiology Res 2014; 3 (01) e8
- 31 Kalaiah MK, Nanchirakal JF, Kharmawphlang L, Noronah SC. Contralateral suppression of transient evoked otoacoustic emissions for various noise signals. Hearing, Balance and Communication 2017; 15 (02) 84-90
- 32 Russo N, Nicol T, Musacchia G, Kraus N. Brainstem responses to speech syllables. Clin Neurophysiol 2004; 115 (09) 2021-2030
- 33 Song JH, Skoe E, Banai K, Kraus N. Perception of speech in noise: neural correlates. J Cogn Neurosci 2011; 23 (09) 2268-2279
- 34 Goodman SS, Mertes IB, Lewis JD, Weissbeck DK. Medial olivocochlear-induced transient-evoked otoacoustic emission amplitude shifts in individual subjects. J Assoc Res Otolaryngol 2013; 14 (06) 829-842
- 35 Jedrzejczak WW, Pilka E, Kochanek K, Skarzynski H. Does the presence of spontaneous components affect the reliability of contralateral suppression of evoked otoacoustic emissions?. Ear Hear 2021; 42 (04) 990-1005