

Subscribe to RSS
DOI: 10.1055/s-0044-1780506
Sequencing, Physiological Regulation, and Representative Disease Research Progress of RNA m6A Modification
Authors
Funding This work is supported by the Huazhong University of Science and Technology Talent Startup Fund; the China Postdoctoral Science Research Fund (Grant No. 217877); and the Open Fund of State Key Laboratory of Natural and Biomimetic Drugs (Peking University) (Grant No. K202218).

Abstract
To date, more than 150 chemical modifications have been disclosed in different RNA species, which are employed to diversify the structure and function of RNA in living organisms. The N 6-methyladenosine (m6A) modification, which is found in the adenosine N 6 site of RNA, has been demonstrated to be the most heavy modification in the mRNA in cells. Moreover, the m6A modification in mRNAs of mammalian and other eukaryotic cells is highly conserved and mandatorily encoded. Increasing evidence indicates that the m6A modification plays a pivotal role in gene-expression regulation and cell-fate decisions. Here, we summarize the most recent m6A-sequencing technology, as well as the molecular mechanism underlying its occurrence, development, and potential use as a target for the treatment of human diseases. Furthermore, our review highlights other newly discovered chemical modifications of RNA that are associated with human disease, as well as their underlying molecular mechanisms. Thus, significant advancements have been made in qualitative/quantitative m6A detection and high-throughput sequencing, and research linking this RNA modification to disease. Efforts toward simplified and more accessible chemical/biological technologies that contribute to precision medicine are ongoing, to benefit society and patients alike.
Introduction
The decades-long development of RNA innovations, from the discovery of the new functions of RNA (serving as a catalyst and regulator of many biochemical reactions) to its conventional role as a genetic-information carrier, linker in protein molecule synthesis, and structural scaffold of subcellular organelles, has contributed to the progress of nucleic acid research.[1] [2] [3] [4] [5] RNA plays an important role in protein synthesis, and RNA-based drug discovery has attracted great interest, as it contains only four types of nucleotides compared with the 20 different amino acid residues present in proteins.[6] To achieve diversity in RNA structure and function, nature uses a variety of chemical groups for its modification.[7] Since the discovery of the foremost RNA modification in yeast about 60 years ago, over 150 distinct modifications have been identified in all RNA species, including transfer RNA (tRNA), messenger RNA (mRNA), and ribosomal RNA (rRNA).[8] [9] [10] [11] These modifications play crucial roles in genetics, growth, and disease development, including embryonic stem cell differentiation, oocyte meiosis, and cancer occurrence.[12] [13] [14] [15] [16] [17]
RNA modifications enrich the diversity of RNA functions and of genetic information processing. An extensive range of RNA modifications has been identified ([Fig. 1]), and the content and related functions of each type vary greatly. Among them, the methylation of RNA nucleotides is the main form of RNA modification, accounting for approximately two-thirds of the total number of RNA modifications. Currently, the methylation modifications found on mRNA include mainly 5-methylcytosine (m5C),[18] 1-methyladenosine (m1A),[19] N 6-methyladenosine (m6A),[20] and pseudouridine (Ψ).[21] Compared with other types of RNA modifications, the m6A internal modification occurs on the N 6 of the adenosine base and is most commonly encountered in eukaryotic mRNA, representing more than 80% of RNA base methylation and having been identified throughout numerous species.[10] [12] [22] [23] [24] [25] [26] [27] It has been reported that m6A can occur in the 5′-untranslated region (UTR), coding sequence (CDS), 3′-UTR, and introns of precursor mRNA, as well as in several noncoding RNAs and miRNAs.[28] [29] In addition to the m6A modification, other hydrophobic modifications of A also occur, such as 2-methylthio-N 6-isopentenyladenosine (ms2i6A),[30] N 6-isopentenyladenosine (i6A),[30] N 6-methyl-threonylcarbamoyl adenosine,[31] and N 6-threonylcarbamoyladenosine (t6A).[32] However, the molecular mechanism underlying these prenyl modifications remains poorly understood ([Fig. 1]; i6A, S-geranylated 2-thiouridines (ges2U)). The biosynthesis mechanism, abundance, and distribution regularity of these prenyl modifications, and the molecular mechanism underlying their regulation network in cells, also remain elusive.

Sequencing Technology Used for m6A Modification
Determining the location of chemical modification groups in transcripts is key information for understanding their basic functions, which is also true for the m6A modification. An analysis of m6A modification over the entire transcriptome indicated that m6A modifications occur across an enormous spectrum of RNA transcripts in unique patterns.[28] [29] In 2012, 30 years after the discovery of the m6A modification, m6A-seq (also known as MeRIP-seq) afforded the m6A landscape in humans and mice. Two independent transcriptome studies have shown that the accurate proportion of m6A-modified RNAs among the total mRNAs is 1 per 2,000 nucleotides, on average.[28] [29] Specifically, total RNA was divided into approximately 100 NTs and subjected to immunoprecipitation using anti-m6A affinity-purified antibodies ([Fig. 2A]). The library prepared from the immunoprecipitated NTs was sequenced, and an algorithm designed to detect peaks was employed to identify the m6A sites.[28] A search of motif databases based on the identified m6A peak sites revealed trends in m6A occurrence in the RRACH consensus sequence (R = G or A; and H = A, C, or U).[28] Although RRACH sequences are ubiquitous in specific transcriptome species, instances of their methylation constitute only 1 to 5% of transcripts in vivo. Intriguingly, the RRACH motif that undergoes m6A methylation is not randomly distributed on transcripts; rather, it occurs in specific CDSs and 3′-untranslated regions (3′-UTRs), and is most commonly observed close to the termination codon. These findings imply that the RRACH motif might not be solely responsible for modulating m6A accumulation.[28] The results obtained in eukaryotic organisms, such as yeast,[33] plants,[34] fruit flies,[35] zebrafish,[36] and mammals,[37] have demonstrated that the cis-regulatory RRACH motif together with CDS/3′-UTR enrichment forms signature traits of the m6A epigenetic transcriptome, thus illustrating the added value of m6A in terms of its function. Given above, the accumulation of m6A in eukaryotic organisms is strictly controlled.

It should be noted that, although m6A-seq can detect m6A modifications at a resolution of 100–200 nt, it lacks information at the single-nucleotide resolution. Site-specific cleavage and radioactive labeling followed by ligation-assisted extraction and thin-layer chromatography (SCARLET) can locate m6A-modification sites down to the individual nucleotide level in RNA samples. Although it is effective for limited applications, it cannot be easily scaled up for the analysis of entire transcriptomes.[38] To boost the granularity of the m6A sequencing information, a technique called photo-crosslinking-assisted m6A sequencing (PA-m6A-seq) has been developed recently. This method uses UV irradiation at 365 nm to crosslink 4-thiouracil with m6A antibodies against mRNA, thereby increasing its resolution to ∼23 nt.[33] miCLIP-seq m6A single-nucleotide resolution crosslinking and immunoprecipitation (CLIP) is another method that can be used in this context, which incorporates the implementation of cutting-edge sequencing strategies for precise genomic characterization. By tracing the distinct alterations at the m6A site that are induced by the UV irradiation of anti-m6A antibodies crosslinked to m6A labeling in RNA, it has been shown that m6A residues often occur in clusters and are frequently distributed in the DRACH (D = A/G/U) motif of CDSs and 3′-UTRs.[39] However, miCLIP requires an input of 20 μg of poly(A)+ mRNA[39] [40]; therefore, it is necessary to optimize its protocols toward a lower input of the initial sample.
The RNA endonuclease MazF from Escherichia coli only cleaves the ACA motif specifically from the 5′ side in the absence of methylation. Based on this discovery, researchers developed a new method termed MAZTER-seq (short for m6A-REF-seq) as an antibody-independent method for analyzing m6A.[41] [42] Although this new method detects m6A methylation only at the ACA site, it identified the enrichment of a distribution pattern near the termination codon, which is consistent with the conclusions drawn from the antibody-based experiments. Furthermore, a sequencing method termed deamination adjacent to RNA modification targets (DART-seq) was developed as an antibody-free m6A-seq method[43] in which the cytosine deaminase apolipoprotein B mRNA editing enzyme catalytic subunit 1 (APOBEC1) is fused to the YT521-B homology (YTH) domain, which binds to m6A to induce C-U deamination at the m6AC sequence, followed by its detection using standard sequencing methods targeting RNA molecules. DART-seq can pinpoint numerous m6A sites, even among small quantities of total RNA from cells (down to 10 ng), and can measure temporal fluctuations in m6A concentrations. However, because effective transfection remains crucial for the performance of the DART-seq method, its usage in living organisms is currently curtailed.[43]
The antibody-free m6A-seq techniques (MAZTER-seq, m6A-REF-seq, and DART-seq) rely on m6A sequencing or cell transfection.[41] [43] A recent study proposed a revolutionary m6A-detection method, i.e., m6A-SEAL, which offers antibody-free fat-mass- and obesity-associated (FTO) protein–assisted chemical labeling for specific detection.[44] In m6A-SEAL, the FTO enzyme promotes the conversion of m6A into a transient hm6A intermediary molecule through thiol addition reactions mediated by DL-dithiothreitol (DTT), thus generating the more stable sulfide-containing dm6A using a simplified installation of biotinylation and other functional tags. The profiling of human and plant transcriptomes using m6A-SEAL confirmed the expected m6A distributions. Moreover, based on comparative analyses against the existing m6A sequencing techniques and the confirmation of specific m6A sites using single-base elongation- and ligation-based qPCR amplification (SELECT), it has been demonstrated that whole-transcriptome scanning via m6A-SEAL yields excellent sensitivity, specificity, and reliability. The versatility of m6A-SEAL, with its FTO-based oxidation and labeling abilities, renders it suitable for diverse applications, ranging from sequencing to enrichment and imaging, thereby advancing m6A research.[44] The combination of this labeling and enrichment strategy with highly specific chemical reactions has attracted great research interest because of its high discrimination ability at the single-base level; furthermore, it has been widely used in studies of the 5-hydroxymethylcytosine, 5-formylcytosine, and pseudouridine (Ψ) RNA modifications.
Biological Function of the M6A Modification: Physiological Regulation and Disease Occurrence
RNA methylation plays an essential role in the regulation of many biological functions, with N 6-methyladenosine being a prominent example for controlling gene expression, translation, and physiology in many organisms, including humans.[45] [46] Three factors govern the reversible m6A modifications ([Fig. 2B]): (1) methyltransferases (Writers), such as methyltransferase-3 (METTL3), methyltransferase-14 (METTL14), and their cofactors, Wilms tumor 1-associated protein (WTAP), KIAA1429 (VIRMA), HAKAI, ZC3H13, RBM15, and RBM15B[47]; (2) m6A-binding proteins (Readers), such as YTH-domain-family proteins and YTH domain-containing protein 1 (YTHDC1)[48]; and (3) demethylases (Erasers), such as the FTO protein and alkylation repair homolog protein 5 (ALKBH5) ([Fig. 3]).[49] [50] Being abundant within eukaryotic cells, m6A controls crucial processes, such as embryonic growth, stem cell specialization, brain-cell generation, and malignancy.

mRNA m6A modulation imparts singular control over transcripts, and its correct deposition in mRNA is crucial for embryonic development.[51] Research has shown that methyltransferase affects the meiosis process in yeast. The fundamental structure of yeast RNA methyltransferase contains three building blocks that form the Mum2–Ime4–Slz1 (MIS) complex: Mum2 (homologous to mammalian WTAP), Ime4 (homologous to mammalian METTL3), and Slz1 (MIS). Mutating any slz1 in yeast does not cause death, but it can lead to impaired meiosis.[52] [53] Arabidopsis thaliana mRNA contains m6A, which is similar to that in animal cells. This modification is necessary for embryogenesis.[54] Inactivation of METLL3 in A. thaliana leads to the failure of continuous transformation throughout the early developmental stages of embryos.[34] [54] During the development of mouse follicles, the ability of RNA metabolism mediated by the m6A methyltransferase KIAA1429 to maintain oocytes is retained.[55] Moreover, m6A determines the transition of endothelial cells to hematopoietic cells during zebrafish embryogenesis, and m6A modification in endothelial cells can specifically regulate the inhibition of the endothelial Notch signals that trigger the emergence of blood-forming progenitors/stem cells. Furthermore, maternal mRNAs that are dependent on m6A are recognized and cleared by YTHDF2, thus promoting the process from zebrafish-fertilized eggs through the maternal-to-zygotic transition.[56] In turn, YTHDC1 guides splicing and polyadenylation decisions during mouse oocyte maturation.[57] Mice lacking YTHDC2 are infertile because their germ cells do not progress past the zygote stage and are essential for sperm production.[58] Absence of ALKBH5 results in higher concentrations of m6A-modified RNAs in male mice, abnormal apoptosis, and reduced fertility.[59] Recently, a transcriptomics investigation identified links between m6A marks in 5′ UTRs and fetal development/pre-eclampsia in the human placenta.[60]
The presence of m6A affects embryonic stem cell maintenance and fate determination.[13] [61] The primordial pluripotent genes of embryonic stem cells, as well as many lineage-specific regulatory genes, carry m6A modifications in their mRNAs.[12] [61] Moreover, the maintenance of stem cell characteristics and destiny relies on proper mRNA m6A modification.[62] The inactivation or depletion of METTL3 in mice and humans leads to prolonged expression of NANOG (which is a transcription factor that is involved in the self-renewal of embryonic stem cells) and delays the turnover of embryonic stem cells caused by self-renewal, thereby preventing their differentiation into downstream lineages in the absence of m6A. This study underscored the significance of m6A for stem cell signaling and homeostasis.[12] Knockout of METTL3 in mice downregulates the m6A levels, causing embryo demise during early gestation.[62] In turn, loss of METTL14 impedes the proliferation and hastens the differentiation of mouse embryonic neural stem cells (NSCs), suggesting that the m6A modification can enhance NSC self-renewal.[63] Recent research indicates that m6A RNA methylation may serve as a promising therapeutic target for various diseases with reduced serum testosterone levels, including azoospermia and oligospermia. A negative correlation exists between the m6A modification and autophagy processes in Leydig cells during testosterone synthesis ([Fig. 4]).[64] These studies demonstrate the significance of m6A during embryonic growth and starting cell difference projects.

m6A and Acute Myeloid Leukemia
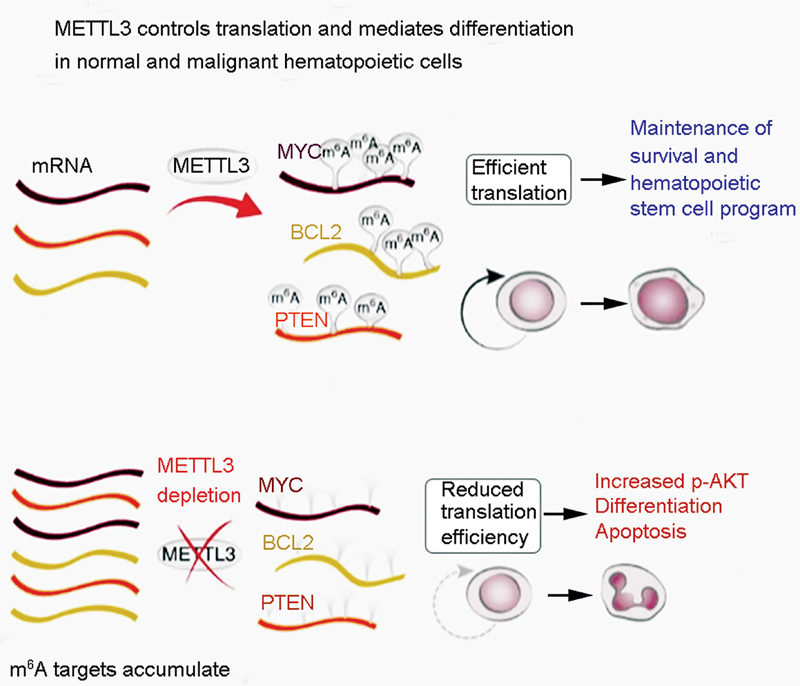
The presence of the m6A modification on mRNA is vital for the advancement and upkeep of acute myeloid leukemia (AML), similar to that observed for the self-restoration of leukemia stem or initiating cells (LSCs/LICs) ([Fig. 5]).[65] Compared with sound hematopoietic undeveloped/progenitor cells (HSPCs) or different types of malignancy cells, the METTL3 quality in AML cells shows an abundance of mRNA and protein expression.[65] The absence of the shaping compound METTL3 in HSPCs advances cell separation and diminishes cell multiplication. METTL3 controls myeloid separation in AML cells through conditional exhaustion, thus prompting cell separation and apoptosis in living beings and postponing the improvement of leukemia in recipient mice.[65] An examination performed at the single-nucleotide level revealed that m6A drives the translation of the cellular myelocytomatosis (c-MYC), B-cell lymphoma 2 (BCL2), and lipid and protein phosphatase and tension homolog (PTEN) mRNAs in a human AML cell line (MOLM-13 cells).[65] Moreover, in AML cells, METTL3 is associated with the CAATT-box binding protein C/EBPZ (CCAAT/enhancer-binding protein zeta) at the transcription initiation site, thus triggering m6A to enhance the translation of related mRNAs, which is crucial for the maintenance of the leukemic state.[11] These investigations provide a hypothetical basis for focusing on METTL3 in AML therapeutically.
m6A and Neurological Diseases
Previous research has confirmed that the m6A modification of mRNA can control neurological outcomes and is important for human brain development and neurological diseases.[66] [67] [68] Studies have found that METTL14 knockout or METTL3 depletion in the cerebrums of developing mouse hatchlings results in the depletion of m6A, thus lengthening the cell cycle and keeping up with the presence of radial nerve cells ([Fig. 6]).[66] The sequencing of m6A in the cerebral mantle of mice revealed an overflow of mRNA particles connected with transcription factors, neurogenesis, the cell cycle, and neuronal separation, whereas m6A marking advanced its disintegration. The presence of m6A also manages human cortical neurogenesis in the organoids of the prefrontal cortex. The examination of the results of m6A-mRNA sequencing during mouse and human cortical neurogenesis revealed a wealth of human-explicit m6A-marked transcripts connected with qualities that favor illnesses of the mind.[66]

Conditional knockout of METTL14 in animals prompted a decline in the number of oligodendrocytes and diminished the myelin layers in the focal sensory system. In vitro, METTL14 depletion can disrupt the maturation of oligodendrocytes after mitosis and has a significant effect on the transcriptomes of precursor cells and oligodendrocytes. In turn, abnormalities of oligodendrocytes can trigger not only demyelinating infections of the focal sensory system, but also neuronal harm or mental issues, and, cerebrum tumors. Moreover, the deletion of METTL14 in oligodendrocyte cell lines induces a strange joining of various RNA molecules.[67] [69] Via behavioral and functional magnetic resonance imaging studies, genetic variations in the m6A demethylase FTO can affect the response of the dopamine-dependent midbrain to reward learning, as well as behavioral responses related to learning from negative outcomes.[70]
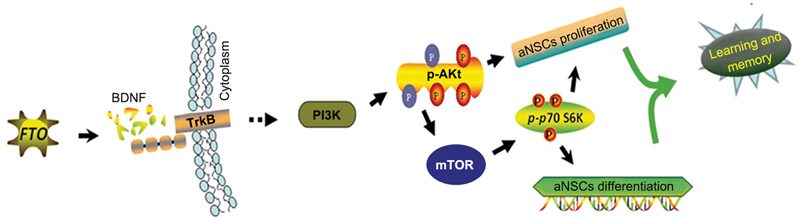
Based on whole-genome m6A analysis and the observation of dynamic m6A modifications during postnatal neurodevelopment, it was determined that FTO deficiency causes changes in the articulation of specific fundamental parts of the brain-prompted neurotrophic factor pathway, denoted by m6A ([Fig. 7]).[71] These examinations suggest that FTO plays a significant role in neurogenesis, learning, and memory. Concomitantly, previous research has found that the m6A demethylase genes FTO and ALKBH5 are associated with major depressive disorder.[72] Furthermore, it has been reported that the insufficiency of FTO can lessen uneasiness and melancholy-like behavior in rodents by changing the gut flora.[73] [74] Moreover, FTO is firmly associated with insulin-inadequacy-related Alzheimer's disease, and conditional deletion of FTO in neurons can decrease intellectual disability in diseased mice.
m6A and Cancers
The m6A modification of mRNA is deeply implicated in the origins and progression of malignancies.[75] [76] The downregulation of the m6A methyltransferase METTL3 or METTL14 significantly promotes the growth, self-renewal, and tumorigenesis of human glioblastoma stem cells (GSCs).[77] [78] METTL3 coordinates the successful execution of carcinogenic pathways in GSCs.[79] An increasing number of studies have confirmed that METTL3 promotes the progression of liver cancer,[80] bladder cancer,[81] gastric cancer ([Fig. 8]),[82] [83] breast cancer ([Fig. 9]),[84] colorectal cancer ([Figs. 10] and [11]),[85] [86] prostate cancer,[87] and other tumors in an m6A-dependent manner.




A recent study confirmed that the miR-186–METTL3 axis promotes the growth of colorectal cancer through the Wnt–β-catenin signaling pathway. The β-catenin signaling pathway promotes the development of hepatoblastoma ([Fig. 12]).[88] In addition, METTL3 can promote chemotherapy and radiation resistance in various tumors, such as pancreatic cancer.[89]

Recently, another study found that the silencing of METTL3 downregulates the mesenchymal–epithelial transition factor (c-Met), phosphorylated Akt (p-Akt), and cell-cycle-related proteins in uveal melanoma (UM) cells, leading to G1 arrest of the cell cycle and restricting UM cell multiplication, aggregation, and mobility.[90] The Epstein–Barr virus (EBV) is a ubiquitous cancer-causing virus that can induce various types of tumors. The reprogramming of EBV antigenic determinants through METTL14 is crucial for the development of virus-associated tumors ([Fig. 13]).[91] The aforementioned studies revealed that the m6A methyltransferase METTL3 or METTL1 may be a potential target for cancer treatment.

The m6A-modified mRNA-binding protein YTHDF1 is overexpressed in various cancers, including non-small-cell lung cancer ([Fig. 14]),[92] colorectal cancer,[93] liver cancer,[94] and ovarian cancer ([Fig. 15])[95]; moreover, the depletion of YTHDF1 can inhibit tumor development. Furthermore, in vitro studies have shown that YTHDF1 knockdown can facilitate the responsiveness to chemotherapy agents, such as fluorouracil and oxaliplatin.[96] Snail is a key transcription factor in the epithelial–mesenchymal transition. Studies of loss and gain of function have confirmed that the YTHDF1-mediated m6A modification of the snail mRNA enhances its translation.[97] Research has demonstrated that by promoting the translation of the m6A-modified cathepsin mRNA, YTHDF1 enhances antigen degradation in phagosomes and limits the cross-presentation of new antigens in dendritic cells, which may become a target for tumor immunotherapy.[98] Moreover, YTHDC2 upregulates HIF-1α and other factors, thus driving tumor spread through the modulation of gene/protein expression.[99] In turn, the inhibition of YTHDC2 can greatly promote the proliferation of tumors, including esophageal squamous cell carcinoma, by affecting several cancer-related signaling pathways.[100] [101]


The insulin-like growth factor-2 mRNA-binding proteins 1, 2, and 3 (IGF2BP1/2/3) are unique families of m6A “readers” that target thousands of mRNA transcripts by recognizing the common GG (m6A) C sequence. These factors may play a carcinogenic role in cancer cells by stabilizing the methylation of mRNAs that target carcinogenic targets, such as c-MYC ([Fig. 16]).[102] [103] [104] In addition, IGF2BP1 can disrupt the stability of the highly upregulated lncRNA in liver cancer through the CCR4–NOT (a highly conserved specific gene silencer) adenylate kinase complex, which is highly linked to liver cancer progression and severity.[105] The IGF2BP3 gene is a downstream target of the carcinogenic effector Lin28b. In a mouse model, it was found that the overexpression of IGF2BP3 drives liver tumorigenesis.[106] Finally, IGF2BP2 and IGF2BP2-2 can promote the occurrence of liver cancer.[107] [108]

It has been reported that the m6A demethylases FTO and ALKBH1 are associated with various malignant tumors,[109] [110] [111] [112] [113] such as the carcinogenic role of FTO in AML ([Fig. 17]).[114] Through autophagy and NF-κ, nutrient-deprivation-triggered FTO in the B pathway leads to heightened melanoma onset in human and murine models. This study suggests that FTO inhibition combined with anti-programmed death-1 (anti-PD-1) blockade may reduce the resistance of melanoma to immunotherapy ([Fig. 18]).[115] By modulating the m6A status of the E2F1 and MYC transcripts, FTO potentiates the proliferation and movement of cervical cancer cells.


Intrahepatic cholangiocarcinoma (ICC) is second only to primary liver cancer as the deadliest form of primary liver cancer with a high prevalence. A Kaplan–Meier survival analysis showed that low expression of FTO promotes the development of ICC, indicating a poor prognosis of ICC.[116] [117] Head and neck squamous cell carcinoma (HNSCC) is the sixth most-common cancer worldwide, with oral squamous cell carcinoma (OSCC) being an aggressive form of HNSCC. Recent research has found that DDX3 (a highly conserved subfamily of the DEAD-box proteins) regulates ALKBH5 to downregulate the m6A methylation level in FOXM1 (Forkhead box, FOX) and NANOG (Nanog Homeobox) new transcripts, thus promoting cisplatin resistance in OSCC.[118] A recent study confirmed that decreased levels of the ALKBH5 and FTO mRNAs are associated with shorter overall survival and cancer-specific survival after nephrectomy.[119] These investigations offer possibilities for novel therapeutic and prognostic avenues for cancer management. The implication of FTO in cancer advancement has prompted interest in drugs such as FB23 and FB23-2, which bind to FTO and inhibit its m6A function.[114] In addition, fluorescein-based compounds have shown potential for targeting FTO demethylation.[120] Meclofenamic acid has been verified as a potent inhibitor that is specific to FTO, with a higher selectivity for FTO versus ALKBH5.[121] The physiological regulation and occurrence of human diseases discussed above focus only on m6A, which is by far the most important chemical modification. In addition to chemical modifications, such as methylation (for example, m6A, m5C, etc.), there is a great need for basic research on the molecular mechanisms of other chemical modifications, together with their links to illness onset and progression in humans, especially the geranyl modification and nicotinamide adenine dinucleotide (NAD) cap modification reported recently.
Recent Research on Newly Discovered RNA Modifications and Their Implications for Cellular Biology
Although the mechanism underlying the RNA chemical modifications remains unclear, especially that of the m6A modification, its role in biological processes has gained increasing recognition, despite the uncertainty about its impact on humans. This review focuses on the progress of high-throughput sequencing technology for the m6A modification; its physiological regulation; and its role in cancer occurrence, cancer development, and tumor suppression. Accumulating evidence indicates that RNA m6A modifiers offer promising biomarker prospects for cancer prognosis and therapy planning, such as in the context of rectal cancer,[122] non-small-cell lung cancer,[123] and renal cell carcinoma.[124] Recent research has found that m6A-mediated long noncoding RNA00958 (LINC00958) upregulation increases adipogenesis and represents a promising nanomedicine target for hepatocellular carcinoma.[125] Because of its pivotal role in multiple diseases, targeting m6A may prove beneficial for the diagnosis and management of illnesses such as AML, glioblastoma, and breast cancer. In addition, scientists recently used the human metapneumovirus (HMPV) as a representative sample, and it was discovered that m6A confers evasion of RNA sensors such as RIG-I on viral RNA (RIG-I), thus confirming that viral RNA m6A can be used as a target for the development of attenuated HMPV vaccines ([Fig. 19]).[126]

In summary, unraveling the significance of and the mechanisms underlying m6A in nucleic acids holds promise for designing new antitumor treatments, which can provide insights for the development of novel therapeutic strategies.
As a popular research field in recent years, the recognition of new types of RNA modification, the investigation of gene activities involved in RNA modifications, and the elucidation of pathogenic mechanisms are valuable avenues for drug development. A typical example is the NAD variant of the RNA cap that commonly occurs outside of the canonical m7G type reported by Liu et al in 2009 ([Fig. 20]). With the advancement of detection technology, NAD cap structures were first discovered in E. coli,[127] followed by yeast, cell lines, plants, and mouse tissues.[128] [129] [130] [131] [132] According to the reported results, NAD cap formation occurs simultaneously with RNA transcription. Concomitantly, the existing RNA sequencing technology has shown that the enzyme NUDT12 (a deNADding enzyme), which removes the NAD cap structure from RNA, has a close regulatory effect on the circadian clock in the liver. Although significant achievements have been reached, e.g., for NAD tagSeq., research on the underlying molecular mechanism remains very limited.

Another typical example was reported in 2012 by Liu et al in which the strongest hydrophobic geranyl (pentenyl dimer) modification to date exists at position 34 of the bacterial tRNA. Geranylation of RNA in vivo is achieved by the enzyme tRNA 2-selenouridine synthase (SelU) ([Fig. 21]). Preliminary studies revealed that the geranyl-modified group can shape the structure and operational properties of RNAs after binding to a specific RNA, as well as its localization within the cell. However, the molecular mechanism underlying this lipid modification is completely unknown.[133] The development of specific markers and localization and enrichment technologies for aryl-modified RNAs will be of great research significance, by laying a solid foundation for explaining its molecular mechanism and even exploring its association with human diseases.

Another noteworthy example is the epoxy-queuosine (oQ) modification reported by Bandarian et al in 2011. The biosynthetic pathway for this modification is shown in [Fig. 22]; however, the molecular mechanism via which the modifying enzymes QueA and QueG regulate the levels of modified oQ and Q remains unknown, especially regarding its possible link to human diseases.[134]

Summary
With the in-depth development of multidisciplinary and interdisciplinary research, an increasing number of new chemically modified RNAs have been reported using comprehensive methods, such as those based on chemistry, life sciences, medicine, and pharmacy. Although these new chemically modified groups perform diverse structural and functional roles in RNA, little is known about their regulatory molecular mechanisms. The development of simple and highly selective chemical and biological methods for their detection, labeling, and localization is of practical significance for elucidating their underlying molecular mechanisms. In particular, the development of highly specific chemical and biological labeling and detection tools for these new chemically modified groups, as well as high-throughput sequencing, will shed light on the molecular underpinnings of RNA modifications. Thus, the development of reliable tools targeting these RNA modifications has great application potential.
In addition, the manner in which these new chemical modification groups are implicated in the pathogenesis of human ailments is particularly noteworthy. The development and application of inhibitors, regulation strategies, and corresponding mRNA-targeted degradation technologies for this new chemically modified enzyme (including encoding and decoding) targets will have practical research significance.
In recent years, in particular, COVID-19 has been rampant worldwide, with very few specific therapeutic drugs or vaccines being available for this condition.[135] Research has shown that there are many unknown chemical modifications in the RNA of this virus that have not been reported. The impact of these modifications on viral characteristics remains elusive, as well as the types, molecular mechanisms, and targets of these modifications.[136] At present, the role played by these new chemical modifications (NAD, prenyl, and oQ modifications) remains unknown,[137] [138] [139] [140] [141] [142] [143] [144] [145] although it has been speculated that they may help the virus avoid host attacks (for example m6A, m1A, and Ψ). Therefore, elucidating the types and characteristics of these chemical modifications is expected to provide new clues for the fight against COVID-19 and many other RNA-relevant viruses. In addition to the application of mature technological tools, the development of new technological toolsets to identify them and the implementation of highly specific small-molecule inhibitors are of great importance to human society.
In summary, significant advancements have been made in qualitative/quantitative m6A detection, high-throughput sequencing, and research linking it to disease. Efforts continue toward the development of simplified and more accessible chemical/biological technologies that will contribute to precision medicine, thus benefiting society and patients alike.
Conflict of Interest
None declared.
Acknowledgments
The authors thank the starting fund from Huazhong University of Science and Technology (HUST) and Shenzhen Huazhong University of Science and Technology Research Institute for financial support.
-
References
- 1 Aas PA, Otterlei M, Falnes PO. et al. Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 2003; 421 (6925): 859-863
- 2 Abbasi-Moheb L, Mertel S, Gonsior M. et al. Mutations in NSUN2 cause autosomal-recessive intellectual disability. Am J Hum Genet 2012; 90 (05) 847-855
- 3 Abeyrathne PD, Koh CS, Grant T, Grigorieff N, Korostelev AA. Ensemble cryo-EM uncovers inchworm-like translocation of a viral IRES through the ribosome. eLife 2016; 5: e14874
- 4 Agris PF, Vendeix FA, Graham WD. tRNA's wobble decoding of the genome: 40 years of modification. J Mol Biol 2007; 366 (01) 1-13
- 5 Anadón C, Guil S, Simó-Riudalbas L. et al. Gene amplification-associated overexpression of the RNA editing enzyme ADAR1 enhances human lung tumorigenesis. Oncogene 2016; 35 (33) 4422
- 6 Appaiah HN, Goswami CP, Mina LA. et al. Persistent upregulation of U6:SNORD44 small RNA ratio in the serum of breast cancer patients. Breast Cancer Res 2011; 13 (05) R86
- 7 Shi H, Wei J, He C. Where, when, and how: context-dependent functions of RNA methylation writers, readers, and erasers. Mol Cell 2019; 74 (04) 640-650
- 8 Balatti V, Nigita G, Veneziano D. et al. tsRNA signatures in cancer. Proc Natl Acad Sci U S A 2017; 114 (30) 8071-8076
- 9 Akichika S, Hirano S, Shichino Y. et al. Cap-specific terminal N 6-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science 2019; 363 (6423): eaav0080
- 10 Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-dependent nuclear RNA processing events. Cell 2015; 162 (06) 1299-1308
- 11 Barbieri I, Tzelepis K, Pandolfini L. et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature 2017; 552 (7683): 126-131
- 12 Batista PJ, Molinie B, Wang J. et al. m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell 2014; 15 (06) 707-719
- 13 Boissel S, Reish O, Proulx K. et al. Loss-of-function mutation in the dioxygenase-encoding FTO gene causes severe growth retardation and multiple malformations. Am J Hum Genet 2009; 85 (01) 106-111
- 14 Roundtree IA, Evans ME, Pan T, He C. Dynamic RNA modifications in gene expression regulation. Cell 2017; 169 (07) 1187-1200
- 15 Sui X, Hu Y, Ren C. et al. METTL3-mediated m6A is required for murine oocyte maturation and maternal-to-zygotic transition. Cell Cycle 2020; 19 (04) 391-404
- 16 Begley U, Sosa MS, Avivar-Valderas A. et al. A human tRNA methyltransferase 9-like protein prevents tumour growth by regulating LIN9 and HIF1-α. EMBO Mol Med 2013; 5 (03) 366-383
- 17 Peng YX, Du J, Wang HS. The roles of m6A in cancer biology and its targeted therapy [in Chinese]. Yao Xue Xue Bao 2019; 54 (10) 1771-1782
- 18 Li Q, Li X, Tang H. et al. NSUN2-Mediated m5C methylation and METTL3/METTL14-mediated m6A methylation cooperatively enhance p21 translation. J Cell Biochem 2017; 118 (09) 2587-2598
- 19 Safra M, Sas-Chen A, Nir R. et al. The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature 2017; 551 (7679): 251-255
- 20 Ries RJ, Zaccara S, Klein P. et al. m6A enhances the phase separation potential of mRNA. Nature 2019; 571 (7765): 424-428
- 21 Zhang W, Eckwahl MJ, Zhou KI, Pan T. Sensitive and quantitative probing of pseudouridine modification in mRNA and long noncoding RNA. RNA 2019; 25 (09) 1218-1225
- 22 Lee Y, Choe J, Park OH, Kim YK. Molecular mechanisms driving mRNA degradation by m6A modification. Trends Genet 2020; 36 (03) 177-188
- 23 Anderson AM, Weasner BP, Weasner BM, Kumar JP. The Drosophila Wilms׳ Tumor 1-Associating Protein (WTAP) homolog is required for eye development. Dev Biol 2014; 390 (02) 170-180
- 24 Bartosovic M, Molares HC, Gregorova P, Hrossova D, Kudla G, Vanacova S. N 6-methyladenosine demethylase FTO targets pre-mRNAs and regulates alternative splicing and 3′-end processing. Nucleic Acids Res 2017; 45 (19) 11356-11370
- 25 Batista PJ. The RNA modification N 6-methyladenosine and its implications in human disease. Genomics Proteomics Bioinformatics 2017; 15 (03) 154-163
- 26 Bertero A, Brown S, Madrigal P. et al. The SMAD2/3 interactome reveals that TGFβ controls m6A mRNA methylation in pluripotency. Nature 2018; 555 (7695): 256-259
- 27 Bodi Z, Zhong S, Mehra S. et al. Adenosine methylation in arabidopsis mRNA is associated with the 3′ end and reduced levels cause developmental defects. Front Plant Sci 2012; 3: 48
- 28 Dominissini D, Moshitch-Moshkovitz S, Schwartz S. et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012; 485 (7397): 201-206
- 29 Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012; 149 (07) 1635-1646
- 30 Schweizer U, Bohleber S, Fradejas-Villar N. The modified base isopentenyladenosine and its derivatives in tRNA. RNA Biol 2017; 14 (09) 1197-1208
- 31 Kimura S, Miyauchi K, Ikeuchi Y, Thiaville PC, Crécy-Lagard Vd, Suzuki T. Discovery of the β-barrel-type RNA methyltransferase responsible for N 6-methylation of N 6-threonylcarbamoyladenosine in tRNAs. Nucleic Acids Res 2014; 42 (14) 9350-9365
- 32 Luthra A, Swinehart W, Bayooz S. et al. Structure and mechanism of a bacterial t6A biosynthesis system. Nucleic Acids Res 2018; 46 (03) 1395-1411
- 33 Schwartz S, Agarwala SD, Mumbach MR. et al. High-resolution mapping reveals a conserved, widespread, dynamic mRNA methylation program in yeast meiosis. Cell 2013; 155 (06) 1409-1421
- 34 Luo GZ, MacQueen A, Zheng G. et al. Unique features of the m6A methylome in Arabidopsis thaliana. Nat Commun 2014; 5: 5630
- 35 Kan L, Grozhik AV, Vedanayagam J. et al. The m6A pathway facilitates sex determination in Drosophila. Nat Commun 2017; 8: 15737
- 36 Zhao BS, Wang X, Beadell AV. et al. m6A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature 2017; 542 (7642): 475-478
- 37 Ma L, Zhao B, Chen K. et al. Evolution of transcript modification by N 6-methyladenosine in primates. Genome Res 2017; 27 (03) 385-392
- 38 Liu N, Parisien M, Dai Q, Zheng G, He C, Pan T. Probing N 6-methyladenosine RNA modification status at single nucleotide resolution in mRNA and long noncoding RNA. RNA 2013; 19 (12) 1848-1856
- 39 Linder B, Grozhik AV, Olarerin-George AO, Meydan C, Mason CE, Jaffrey SR. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods 2015; 12 (08) 767-772
- 40 Imanishi M, Tsuji S, Suda A, Futaki S. Detection of N 6-methyladenosine based on the methyl-sensitivity of MazF RNA endonuclease. Chem Commun (Camb) 2017; 53 (96) 12930-12933
- 41 Garcia-Campos MA, Edelheit S, Toth U. et al. Deciphering the “m6A Code” via antibody-independent quantitative profiling. Cell 2019; 178 (03) 731-747.e16
- 42 Zhang Z, Chen LQ, Zhao YL. et al. Single-base mapping of m6A by an antibody-independent method. Sci Adv 2019; 5 (07) eaax0250
- 43 Meyer KD. DART-seq: an antibody-free method for global m6A detection. Nat Methods 2019; 16 (12) 1275-1280
- 44 Wang Y, Xiao Y, Dong S, Yu Q, Jia G. Antibody-free enzyme-assisted chemical approach for detection of N 6-methyladenosine. Nat Chem Biol 2020; 16 (08) 896-903
- 45 Coker H, Wei G, Brockdorff N. m6A modification of non-coding RNA and the control of mammalian gene expression. Biochim Biophys Acta Gene Regul Mech 2019; 1862 (03) 310-318
- 46 Lan Q, Liu PY, Haase J, Bell JL, Hüttelmaier S, Liu T. The critical role of RNA m6A methylation in cancer. Cancer Res 2019; 79 (07) 1285-1292
- 47 Du Y, Hou G, Zhang H. et al. SUMOylation of the m6A-RNA methyltransferase METTL3 modulates its function. Nucleic Acids Res 2018; 46 (10) 5195-5208
- 48 Ho AJ, Stein JL, Hua X. et al; Alzheimer's Disease Neuroimaging Initiative. A commonly carried allele of the obesity-related FTO gene is associated with reduced brain volume in the healthy elderly. Proc Natl Acad Sci U S A 2010; 107 (18) 8404-8409
- 49 Li J, Han Y, Zhang H. et al. The m6A demethylase FTO promotes the growth of lung cancer cells by regulating the m6A level of USP7 mRNA. Biochem Biophys Res Commun 2019; 512 (03) 479-485
- 50 Li XC, Jin F, Wang BY, Yin XJ, Hong W, Tian FJ. The m6A demethylase ALKBH5 controls trophoblast invasion at the maternal-fetal interface by regulating the stability of CYR61 mRNA. Theranostics 2019; 9 (13) 3853-3865
- 51 Fazi F, Fatica A. Interplay between N 6-methyladenosine (m6A) and non-coding RNAs in cell development and cancer. Front Cell Dev Biol 2019; 7: 116
- 52 Agarwala SD, Blitzblau HG, Hochwagen A, Fink GR. RNA methylation by the MIS complex regulates a cell fate decision in yeast. PLoS Genet 2012; 8 (06) e1002732
- 53 Hongay CF, Grisafi PL, Galitski T, Fink GR. Antisense transcription controls cell fate in Saccharomyces cerevisiae. Cell 2006; 127 (04) 735-745
- 54 Reichel M, Köster T, Staiger D. Marking RNA: m6A writers, readers, and functions in arabidopsis. J Mol Cell Biol 2019; 11 (10) 899-910
- 55 Hu Y, Ouyang Z, Sui X. et al. Oocyte competence is maintained by m6A methyltransferase KIAA1429-mediated RNA metabolism during mouse follicular development. Cell Death Differ 2020; 27 (08) 2468-2483
- 56 Hsu PJ, He C. Making changes: N 6-methyladenosine-mediated decay drives the endothelial-to-hematopoietic transition. Biochemistry 2017; 56 (46) 6077-6078
- 57 Kasowitz SD, Ma J, Anderson SJ. et al. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLoS Genet 2018; 14 (05) e1007412
- 58 Hsu PJ, Zhu Y, Ma H. et al. Ythdc2 is an N 6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res 2017; 27 (09) 1115-1127
- 59 Zheng G, Dahl JA, Niu Y. et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell 2013; 49 (01) 18-29
- 60 Taniguchi K, Kawai T, Kitawaki J. et al. Epitranscriptomic profiling in human placenta: N 6-methyladenosine modification at the 5′-untranslated region is related to fetal growth and preeclampsia. FASEB J 2020; 34 (01) 494-512
- 61 Wang Y, Li Y, Toth JI, Petroski MD, Zhang Z, Zhao JC. N 6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat Cell Biol 2014; 16 (02) 191-198
- 62 Geula S, Moshitch-Moshkovitz S, Dominissini D. et al. Stem cells. m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science 2015; 347 (6225): 1002-1006
- 63 Wang Y, Li Y, Yue M. et al. N 6-methyladenosine RNA modification regulates embryonic neural stem cell self-renewal through histone modifications. Nat Neurosci 2018; 21 (02) 195-206
- 64 Chen Y, Wang J, Xu D. et al. m6A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy 2021; 17 (02) 457-475
- 65 Vu LP, Pickering BF, Cheng Y. et al. The N 6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat Med 2017; 23 (11) 1369-1376
- 66 Yoon KJ, Ringeling FR, Vissers C. et al. Temporal control of mammalian cortical neurogenesis by m6A methylation. Cell 2017; 171 (04) 877-889.e17
- 67 Xu H, Dzhashiashvili Y, Shah A. et al. m6A mRNA methylation is essential for oligodendrocyte maturation and CNS myelination. Neuron 2020; 105 (02) 293.e5-309.e5
- 68 Livneh I, Moshitch-Moshkovitz S, Amariglio N, Rechavi G, Dominissini D. The m6A epitranscriptome: transcriptome plasticity in brain development and function. Nat Rev Neurosci 2020; 21 (01) 36-51
- 69 Koreman E, Sun X, Lu QR. Chromatin remodeling and epigenetic regulation of oligodendrocyte myelination and myelin repair. Mol Cell Neurosci 2018; 87: 18-26
- 70 Sevgi M, Rigoux L, Kühn AB. et al. An obesity-predisposing variant of the FTO gene regulates D2R-dependent reward learning. J Neurosci 2015; 35 (36) 12584-12592
- 71 Li L, Zang L, Zhang F. et al. Fat mass and obesity-associated (FTO) protein regulates adult neurogenesis. Hum Mol Genet 2017; 26 (13) 2398-2411
- 72 Du T, Rao S, Wu L. et al. An association study of the m6A genes with major depressive disorder in Chinese Han population. J Affect Disord 2015; 183: 279-286
- 73 Sun L, Ma L, Zhang H. et al. Fto deficiency reduces anxiety- and depression-like behaviors in mice via alterations in gut microbiota. Theranostics 2019; 9 (03) 721-733
- 74 Li H, Ren Y, Mao K. et al. FTO is involved in Alzheimer's disease by targeting TSC1-mTOR-Tau signaling. Biochem Biophys Res Commun 2018; 498 (01) 234-239
- 75 Ma S, Chen C, Ji X. et al. The interplay between m6A RNA methylation and noncoding RNA in cancer. J Hematol Oncol 2019; 12 (01) 121
- 76 He L, Li H, Wu A, Peng Y, Shu G, Yin G. Functions of N 6-methyladenosine and its role in cancer. Mol Cancer 2019; 18 (01) 176
- 77 Cui Q, Shi H, Ye P. et al. m6A RNA methylation regulates the self-renewal and tumorigenesis of glioblastoma stem cells. Cell Rep 2017; 18 (11) 2622-2634
- 78 Visvanathan A, Patil V, Arora A. et al. Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2018; 37 (04) 522-533
- 79 Visvanathan A, Patil V, Abdulla S, Hoheisel JD, Somasundaram K. N 6-methyladenosine landscape of glioma stem-like cells: METTL3 is essential for the expression of actively transcribed genes and sustenance of the oncogenic signaling. Genes (Basel) 2019; 10 (02) 141
- 80 Chen M, Wei L, Law CT. et al. RNA N 6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018; 67 (06) 2254-2270
- 81 Han J, Wang JZ, Yang X. et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol Cancer 2019; 18 (01) 110
- 82 Wang Q, Chen C, Ding Q. et al. METTL3-mediated m6A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut 2020; 69 (07) 1193-1205
- 83 Lin S, Liu J, Jiang W. et al. METTL3 promotes the proliferation and mobility of gastric cancer cells. Open Med (Wars) 2019; 14: 25-31
- 84 Cai X, Wang X, Cao C. et al. HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tumor suppressor let-7g. Cancer Lett 2018; 415: 11-19
- 85 Li T, Hu PS, Zuo Z. et al. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer 2019; 18 (01) 112
- 86 Zhu W, Si Y, Xu J. et al. Methyltransferase like 3 promotes colorectal cancer proliferation by stabilizing CCNE1 mRNA in an m6A-dependent manner. J Cell Mol Med 2020; 24 (06) 3521-3533
- 87 Cai J, Yang F, Zhan H. et al. RNA m6A methyltransferase METTL3 promotes the growth of prostate cancer by regulating hedgehog pathway. OncoTargets Ther 2019; 12: 9143-9152
- 88 Cui X, Wang Z, Li J. et al. Cross talk between RNA N 6-methyladenosine methyltransferase-like 3 and miR-186 regulates hepatoblastoma progression through Wnt/β-catenin signalling pathway. Cell Prolif 2020; 53 (03) e12768
- 89 Taketo K, Konno M, Asai A. et al. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int J Oncol 2018; 52 (02) 621-629
- 90 Luo G, Xu W, Zhao Y. et al. RNA m6 A methylation regulates uveal melanoma cell proliferation, migration, and invasion by targeting c-Met. J Cell Physiol 2020; 235 (10) 7107-7119
- 91 Lang F, Singh RK, Pei Y, Zhang S, Sun K, Robertson ES. EBV epitranscriptome reprogramming by METTL14 is critical for viral-associated tumorigenesis. PLoS Pathog 2019; 15 (06) e1007796
- 92 Shi Y, Fan S, Wu M. et al. YTHDF1 links hypoxia adaptation and non-small cell lung cancer progression. Nat Commun 2019; 10 (01) 4892
- 93 Bai Y, Yang C, Wu R. et al. YTHDF1 regulates tumorigenicity and cancer stem cell-like activity in human colorectal carcinoma. Front Oncol 2019; 9: 332
- 94 Zhao X, Chen Y, Mao Q. et al. Overexpression of YTHDF1 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Biomark 2018; 21 (04) 859-868
- 95 Liu T, Wei Q, Jin J. et al. The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation. Nucleic Acids Res 2020; 48 (07) 3816-3831
- 96 Nishizawa Y, Konno M, Asai A. et al. Oncogene c-Myc promotes epitranscriptome m6A reader YTHDF1 expression in colorectal cancer. Oncotarget 2017; 9 (07) 7476-7486
- 97 Lin X, Chai G, Wu Y. et al. RNA m6A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail. Nat Commun 2019; 10 (01) 2065
- 98 Kim DJ, Iwasaki A. YTHDF1 control of dendritic cell cross-priming as a possible target of cancer immunotherapy. Biochemistry 2019; 58 (15) 1945-1946
- 99 Tanabe A, Tanikawa K, Tsunetomi M. et al. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett 2016; 376 (01) 34-42
- 100 Yang N, Ying P, Tian J. et al. Genetic variants in m6A modification genes are associated with esophageal squamous-cell carcinoma in the Chinese population. Carcinogenesis 2020; 41 (06) 761-768
- 101 Fanale D, Iovanna JL, Calvo EL. et al. Germline copy number variation in the YTHDC2 gene: does it have a role in finding a novel potential molecular target involved in pancreatic adenocarcinoma susceptibility?. Expert Opin Ther Targets 2014; 18 (08) 841-850
- 102 Huang H, Weng H, Sun W. et al. Recognition of RNA N 6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol 2018; 20 (03) 285-295
- 103 Gutschner T, Hämmerle M, Pazaitis N. et al. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) is an important protumorigenic factor in hepatocellular carcinoma. Hepatology 2014; 59 (05) 1900-1911
- 104 Müller S, Glaß M, Singh AK. et al. IGF2BP1 promotes SRF-dependent transcription in cancer in a m6A- and miRNA-dependent manner. Nucleic Acids Res 2019; 47 (01) 375-390
- 105 Hämmerle M, Gutschner T, Uckelmann H. et al. Posttranscriptional destabilization of the liver-specific long noncoding RNA HULC by the IGF2 mRNA-binding protein 1 (IGF2BP1). Hepatology 2013; 58 (05) 1703-1712
- 106 Nguyen LH, Robinton DA, Seligson MT. et al. Lin28b is sufficient to drive liver cancer and necessary for its maintenance in murine models. Cancer Cell 2014; 26 (02) 248-261
- 107 Dai N, Zhao L, Wrighting D. et al. IGF2BP2/IMP2-deficient mice resist obesity through enhanced translation of Ucp1 mRNA and other mRNAs encoding mitochondrial proteins. Cell Metab 2015; 21 (04) 609-621
- 108 Simon Y, Kessler SM, Bohle RM, Haybaeck J, Kiemer AK. The insulin-like growth factor 2 (IGF2) mRNA-binding protein p62/IGF2BP2-2 as a promoter of NAFLD and HCC?. Gut 2014; 63 (05) 861-863
- 109 Xu D, Shao W, Jiang Y, Wang X, Liu Y, Liu X. FTO expression is associated with the occurrence of gastric cancer and prognosis. Oncol Rep 2017; 38 (04) 2285-2292
- 110 Deng X, Su R, Stanford S, Chen J. Critical enzymatic functions of FTO in obesity and cancer. Front Endocrinol (Lausanne) 2018; 9: 396
- 111 Chen J, Du B. Novel positioning from obesity to cancer: FTO, an m6A RNA demethylase, regulates tumour progression. J Cancer Res Clin Oncol 2019; 145 (01) 19-29
- 112 Li Y, Zheng D, Wang F, Xu Y, Yu H, Zhang H. Expression of demethylase genes, FTO and ALKBH1, is associated with prognosis of gastric cancer. Dig Dis Sci 2019; 64 (06) 1503-1513
- 113 Tang B, Yang Y, Kang M. et al. m6A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling. Mol Cancer 2020; 19 (01) 3
- 114 Huang Y, Su R, Sheng Y. et al. Small-molecule targeting of oncogenic FTO demethylase in acute myeloid leukemia. Cancer Cell 2019; 35 (04) 677-691.e10
- 115 Yang S, Wei J, Cui YH. et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat Commun 2019; 10 (01) 2782
- 116 Zou D, Dong L, Li C, Yin Z, Rao S, Zhou Q. The m6A eraser FTO facilitates proliferation and migration of human cervical cancer cells. Cancer Cell Int 2019; 19: 321
- 117 Rong ZX, Li Z, He JJ. et al. Downregulation of fat mass and obesity associated (FTO) promotes the progression of intrahepatic cholangiocarcinoma. Front Oncol 2019; 9: 369
- 118 Shriwas O, Priyadarshini M, Samal SK. et al. DDX3 modulates cisplatin resistance in OSCC through ALKBH5-mediated m6A-demethylation of FOXM1 and NANOG. Apoptosis 2020; 25 (3–4): 233-246
- 119 Strick A, von Hagen F, Gundert L. et al. The N 6 -methyladenosine (m6 A) erasers alkylation repair homologue 5 (ALKBH5) and fat mass and obesity-associated protein (FTO) are prognostic biomarkers in patients with clear cell renal carcinoma. BJU Int 2020; 125 (04) 617-624
- 120 Wang T, Hong T, Huang Y. et al. Fluorescein derivatives as bifunctional molecules for the simultaneous inhibiting and labeling of FTO protein. J Am Chem Soc 2015; 137 (43) 13736-13739
- 121 Huang Y, Yan J, Li Q. et al. Meclofenamic acid selectively inhibits FTO demethylation of m6A over ALKBH5. Nucleic Acids Res 2015; 43 (01) 373-384
- 122 Zhuang J, Lin C, Ye J. m6 A RNA methylation regulators contribute to malignant progression in rectal cancer. J Cell Physiol 2020; 235 (09) 6300-6306
- 123 Liu Y, Guo X, Zhao M. et al. Contributions and prognostic values of m6 A RNA methylation regulators in non-small-cell lung cancer. J Cell Physiol 2020; 235 (09) 6043-6057
- 124 Wang J, Zhang C, He W, Gou X. Effect of m6A RNA methylation regulators on malignant progression and prognosis in renal clear cell carcinoma. Front Oncol 2020; 10: 3
- 125 Zuo X, Chen Z, Gao W. et al. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J Hematol Oncol 2020; 13 (01) 5
- 126 Lu M, Zhang Z, Xue M. et al. N 6-methyladenosine modification enables viral RNA to escape recognition by RNA sensor RIG-I. Nat Microbiol 2020; 5 (04) 584-598
- 127 Chen YG, Kowtoniuk WE, Agarwal I, Shen Y, Liu DR. LC/MS analysis of cellular RNA reveals NAD-linked RNA. Nat Chem Biol 2009; 5 (12) 879-881
- 128 Cahová H, Winz ML, Höfer K, Nübel G, Jäschke A. NAD captureSeq indicates NAD as a bacterial cap for a subset of regulatory RNAs. Nature 2015; 519 (7543): 374-377
- 129 Walters RW, Matheny T, Mizoue LS, Rao BS, Muhlrad D, Parker R. Identification of NAD+ capped mRNAs in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 2017; 114 (03) 480-485
- 130 Jiao X, Doamekpor SK, Bird JG. et al. 5′ End nicotinamide adenine dinucleotide cap in human cells promotes RNA decay through DXO-mediated deNADding. Cell 2017; 168 (06) 1015-1027.e10
- 131 Zhang H, Zhong H, Zhang S. et al. NAD tagSeq reveals that NAD+-capped RNAs are mostly produced from a large number of protein-coding genes in Arabidopsis . Proc Natl Acad Sci U S A 2019; 116 (24) 12072-12077
- 132 Bird JG, Basu U, Kuster D. et al. Highly efficient 5′ capping of mitochondrial RNA with NAD+ and NADH by yeast and human mitochondrial RNA polymerase. eLife 2018; 7: e42179
- 133 Dumelin CE, Chen Y, Leconte AM, Chen YG, Liu DR. Discovery and biological characterization of geranylated RNA in bacteria. Nat Chem Biol 2012; 8 (11) 913-919
- 134 Miles ZD, McCarty RM, Molnar G, Bandarian V. Discovery of epoxyqueuosine (oQ) reductase reveals parallels between halorespiration and tRNA modification. Proc Natl Acad Sci U S A 2011; 108 (18) 7368-7372
- 135 Li J, Jiang XY, Xu SJ. et al. Medicinal chemistry strategies in seeking coronavirus inhibitors. Yao Xue Xue Bao 2020; 55 (04) 537-553
- 136 Kim D, Lee JY, Yang JS, Kim JW, Kim VN, Chang H. The architecture of SARS-CoV-2 transcriptome. Cell 2020; 181 (04) 914-921.e10
- 137 Zhou H, Gan Y, Li Y, Chen X, Guo Y, Wang R. Degradation of rat sarcoma proteins targeting the post-translational prenyl modifications via cascade azidation/fluorination and click reaction. J Med Chem 2023; 66 (11) 7243-7252
- 138 Gan Y, Li Y, Zhou H, Wang R. Deciphering regulatory proteins of prenylated protein via the FRET technique using nitroso-based ene-ligation and sequential azidation and click reaction. Org Lett 2022; 24 (36) 6625-6630
- 139 Gan Y, Chen X, Li Y, Guo Y, Wang R. Sequential azidation/azolation of prenylated derivatives and a click reaction enable selective labeling and degradation of RAS protein. J Org Chem 2023; 88 (15) 10836-10843
- 140 Li Y, Zhou H, Liu H. et al. Development of nitroso-based probes for labeling and regulation of RAS proteins in cancer cells via sequential ene-ligation and oxime condensation. J Org Chem 2023; 88 (03) 1762-1771
- 141 Wang S, Li Y, Zhou H, Wang L, Wang R. Development of biocompatible ene-ligation enabled by prenyl-based β-caryophyllene with triazoline/selectfluor under physiological conditions. J Org Chem 2022; 87 (13) 8648-8655
- 142 Zhou H, Li Y, Gan Y, Wang R. Total RNA synthesis and its covalent labeling innovation. Top Curr Chem (Cham) 2022; 380 (03) 16
- 143 Gan YF, Li YY, Chen XQ, Guo YY, Wang R. Recent advances in quinone methide chemistry for protein-proximity capturing. Synthesis 2023; 55 (08) 1172-1186
- 144 Wang R, Vangaveti S, Ranganathan SV. et al. Synthesis, base pairing and structure studies of geranylated RNA. Nucleic Acids Res 2016; 44 (13) 6036-6045
- 145 Li Y, Zhou H, Chen S. et al. Bioorthogonal labeling and profiling of N 6-isopentenyladenosine (i6A) modified RNA. Nucleic Acids Res 2024 (e-pub ahead of print).
Address for correspondence
Publication History
Received: 24 October 2023
Accepted: 24 January 2024
Article published online:
11 March 2024
© 2024. The Author(s). This is an open access article published by Thieme under the terms of the Creative Commons Attribution License, permitting unrestricted use, distribution, and reproduction so long as the original work is properly cited. (https://creativecommons.org/licenses/by/4.0/)
Georg Thieme Verlag KG
Rüdigerstraße 14, 70469 Stuttgart, Germany
-
References
- 1 Aas PA, Otterlei M, Falnes PO. et al. Human and bacterial oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 2003; 421 (6925): 859-863
- 2 Abbasi-Moheb L, Mertel S, Gonsior M. et al. Mutations in NSUN2 cause autosomal-recessive intellectual disability. Am J Hum Genet 2012; 90 (05) 847-855
- 3 Abeyrathne PD, Koh CS, Grant T, Grigorieff N, Korostelev AA. Ensemble cryo-EM uncovers inchworm-like translocation of a viral IRES through the ribosome. eLife 2016; 5: e14874
- 4 Agris PF, Vendeix FA, Graham WD. tRNA's wobble decoding of the genome: 40 years of modification. J Mol Biol 2007; 366 (01) 1-13
- 5 Anadón C, Guil S, Simó-Riudalbas L. et al. Gene amplification-associated overexpression of the RNA editing enzyme ADAR1 enhances human lung tumorigenesis. Oncogene 2016; 35 (33) 4422
- 6 Appaiah HN, Goswami CP, Mina LA. et al. Persistent upregulation of U6:SNORD44 small RNA ratio in the serum of breast cancer patients. Breast Cancer Res 2011; 13 (05) R86
- 7 Shi H, Wei J, He C. Where, when, and how: context-dependent functions of RNA methylation writers, readers, and erasers. Mol Cell 2019; 74 (04) 640-650
- 8 Balatti V, Nigita G, Veneziano D. et al. tsRNA signatures in cancer. Proc Natl Acad Sci U S A 2017; 114 (30) 8071-8076
- 9 Akichika S, Hirano S, Shichino Y. et al. Cap-specific terminal N 6-methylation of RNA by an RNA polymerase II-associated methyltransferase. Science 2019; 363 (6423): eaav0080
- 10 Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-dependent nuclear RNA processing events. Cell 2015; 162 (06) 1299-1308
- 11 Barbieri I, Tzelepis K, Pandolfini L. et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature 2017; 552 (7683): 126-131
- 12 Batista PJ, Molinie B, Wang J. et al. m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell 2014; 15 (06) 707-719
- 13 Boissel S, Reish O, Proulx K. et al. Loss-of-function mutation in the dioxygenase-encoding FTO gene causes severe growth retardation and multiple malformations. Am J Hum Genet 2009; 85 (01) 106-111
- 14 Roundtree IA, Evans ME, Pan T, He C. Dynamic RNA modifications in gene expression regulation. Cell 2017; 169 (07) 1187-1200
- 15 Sui X, Hu Y, Ren C. et al. METTL3-mediated m6A is required for murine oocyte maturation and maternal-to-zygotic transition. Cell Cycle 2020; 19 (04) 391-404
- 16 Begley U, Sosa MS, Avivar-Valderas A. et al. A human tRNA methyltransferase 9-like protein prevents tumour growth by regulating LIN9 and HIF1-α. EMBO Mol Med 2013; 5 (03) 366-383
- 17 Peng YX, Du J, Wang HS. The roles of m6A in cancer biology and its targeted therapy [in Chinese]. Yao Xue Xue Bao 2019; 54 (10) 1771-1782
- 18 Li Q, Li X, Tang H. et al. NSUN2-Mediated m5C methylation and METTL3/METTL14-mediated m6A methylation cooperatively enhance p21 translation. J Cell Biochem 2017; 118 (09) 2587-2598
- 19 Safra M, Sas-Chen A, Nir R. et al. The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature 2017; 551 (7679): 251-255
- 20 Ries RJ, Zaccara S, Klein P. et al. m6A enhances the phase separation potential of mRNA. Nature 2019; 571 (7765): 424-428
- 21 Zhang W, Eckwahl MJ, Zhou KI, Pan T. Sensitive and quantitative probing of pseudouridine modification in mRNA and long noncoding RNA. RNA 2019; 25 (09) 1218-1225
- 22 Lee Y, Choe J, Park OH, Kim YK. Molecular mechanisms driving mRNA degradation by m6A modification. Trends Genet 2020; 36 (03) 177-188
- 23 Anderson AM, Weasner BP, Weasner BM, Kumar JP. The Drosophila Wilms׳ Tumor 1-Associating Protein (WTAP) homolog is required for eye development. Dev Biol 2014; 390 (02) 170-180
- 24 Bartosovic M, Molares HC, Gregorova P, Hrossova D, Kudla G, Vanacova S. N 6-methyladenosine demethylase FTO targets pre-mRNAs and regulates alternative splicing and 3′-end processing. Nucleic Acids Res 2017; 45 (19) 11356-11370
- 25 Batista PJ. The RNA modification N 6-methyladenosine and its implications in human disease. Genomics Proteomics Bioinformatics 2017; 15 (03) 154-163
- 26 Bertero A, Brown S, Madrigal P. et al. The SMAD2/3 interactome reveals that TGFβ controls m6A mRNA methylation in pluripotency. Nature 2018; 555 (7695): 256-259
- 27 Bodi Z, Zhong S, Mehra S. et al. Adenosine methylation in arabidopsis mRNA is associated with the 3′ end and reduced levels cause developmental defects. Front Plant Sci 2012; 3: 48
- 28 Dominissini D, Moshitch-Moshkovitz S, Schwartz S. et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012; 485 (7397): 201-206
- 29 Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell 2012; 149 (07) 1635-1646
- 30 Schweizer U, Bohleber S, Fradejas-Villar N. The modified base isopentenyladenosine and its derivatives in tRNA. RNA Biol 2017; 14 (09) 1197-1208
- 31 Kimura S, Miyauchi K, Ikeuchi Y, Thiaville PC, Crécy-Lagard Vd, Suzuki T. Discovery of the β-barrel-type RNA methyltransferase responsible for N 6-methylation of N 6-threonylcarbamoyladenosine in tRNAs. Nucleic Acids Res 2014; 42 (14) 9350-9365
- 32 Luthra A, Swinehart W, Bayooz S. et al. Structure and mechanism of a bacterial t6A biosynthesis system. Nucleic Acids Res 2018; 46 (03) 1395-1411
- 33 Schwartz S, Agarwala SD, Mumbach MR. et al. High-resolution mapping reveals a conserved, widespread, dynamic mRNA methylation program in yeast meiosis. Cell 2013; 155 (06) 1409-1421
- 34 Luo GZ, MacQueen A, Zheng G. et al. Unique features of the m6A methylome in Arabidopsis thaliana. Nat Commun 2014; 5: 5630
- 35 Kan L, Grozhik AV, Vedanayagam J. et al. The m6A pathway facilitates sex determination in Drosophila. Nat Commun 2017; 8: 15737
- 36 Zhao BS, Wang X, Beadell AV. et al. m6A-dependent maternal mRNA clearance facilitates zebrafish maternal-to-zygotic transition. Nature 2017; 542 (7642): 475-478
- 37 Ma L, Zhao B, Chen K. et al. Evolution of transcript modification by N 6-methyladenosine in primates. Genome Res 2017; 27 (03) 385-392
- 38 Liu N, Parisien M, Dai Q, Zheng G, He C, Pan T. Probing N 6-methyladenosine RNA modification status at single nucleotide resolution in mRNA and long noncoding RNA. RNA 2013; 19 (12) 1848-1856
- 39 Linder B, Grozhik AV, Olarerin-George AO, Meydan C, Mason CE, Jaffrey SR. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods 2015; 12 (08) 767-772
- 40 Imanishi M, Tsuji S, Suda A, Futaki S. Detection of N 6-methyladenosine based on the methyl-sensitivity of MazF RNA endonuclease. Chem Commun (Camb) 2017; 53 (96) 12930-12933
- 41 Garcia-Campos MA, Edelheit S, Toth U. et al. Deciphering the “m6A Code” via antibody-independent quantitative profiling. Cell 2019; 178 (03) 731-747.e16
- 42 Zhang Z, Chen LQ, Zhao YL. et al. Single-base mapping of m6A by an antibody-independent method. Sci Adv 2019; 5 (07) eaax0250
- 43 Meyer KD. DART-seq: an antibody-free method for global m6A detection. Nat Methods 2019; 16 (12) 1275-1280
- 44 Wang Y, Xiao Y, Dong S, Yu Q, Jia G. Antibody-free enzyme-assisted chemical approach for detection of N 6-methyladenosine. Nat Chem Biol 2020; 16 (08) 896-903
- 45 Coker H, Wei G, Brockdorff N. m6A modification of non-coding RNA and the control of mammalian gene expression. Biochim Biophys Acta Gene Regul Mech 2019; 1862 (03) 310-318
- 46 Lan Q, Liu PY, Haase J, Bell JL, Hüttelmaier S, Liu T. The critical role of RNA m6A methylation in cancer. Cancer Res 2019; 79 (07) 1285-1292
- 47 Du Y, Hou G, Zhang H. et al. SUMOylation of the m6A-RNA methyltransferase METTL3 modulates its function. Nucleic Acids Res 2018; 46 (10) 5195-5208
- 48 Ho AJ, Stein JL, Hua X. et al; Alzheimer's Disease Neuroimaging Initiative. A commonly carried allele of the obesity-related FTO gene is associated with reduced brain volume in the healthy elderly. Proc Natl Acad Sci U S A 2010; 107 (18) 8404-8409
- 49 Li J, Han Y, Zhang H. et al. The m6A demethylase FTO promotes the growth of lung cancer cells by regulating the m6A level of USP7 mRNA. Biochem Biophys Res Commun 2019; 512 (03) 479-485
- 50 Li XC, Jin F, Wang BY, Yin XJ, Hong W, Tian FJ. The m6A demethylase ALKBH5 controls trophoblast invasion at the maternal-fetal interface by regulating the stability of CYR61 mRNA. Theranostics 2019; 9 (13) 3853-3865
- 51 Fazi F, Fatica A. Interplay between N 6-methyladenosine (m6A) and non-coding RNAs in cell development and cancer. Front Cell Dev Biol 2019; 7: 116
- 52 Agarwala SD, Blitzblau HG, Hochwagen A, Fink GR. RNA methylation by the MIS complex regulates a cell fate decision in yeast. PLoS Genet 2012; 8 (06) e1002732
- 53 Hongay CF, Grisafi PL, Galitski T, Fink GR. Antisense transcription controls cell fate in Saccharomyces cerevisiae. Cell 2006; 127 (04) 735-745
- 54 Reichel M, Köster T, Staiger D. Marking RNA: m6A writers, readers, and functions in arabidopsis. J Mol Cell Biol 2019; 11 (10) 899-910
- 55 Hu Y, Ouyang Z, Sui X. et al. Oocyte competence is maintained by m6A methyltransferase KIAA1429-mediated RNA metabolism during mouse follicular development. Cell Death Differ 2020; 27 (08) 2468-2483
- 56 Hsu PJ, He C. Making changes: N 6-methyladenosine-mediated decay drives the endothelial-to-hematopoietic transition. Biochemistry 2017; 56 (46) 6077-6078
- 57 Kasowitz SD, Ma J, Anderson SJ. et al. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLoS Genet 2018; 14 (05) e1007412
- 58 Hsu PJ, Zhu Y, Ma H. et al. Ythdc2 is an N 6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res 2017; 27 (09) 1115-1127
- 59 Zheng G, Dahl JA, Niu Y. et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell 2013; 49 (01) 18-29
- 60 Taniguchi K, Kawai T, Kitawaki J. et al. Epitranscriptomic profiling in human placenta: N 6-methyladenosine modification at the 5′-untranslated region is related to fetal growth and preeclampsia. FASEB J 2020; 34 (01) 494-512
- 61 Wang Y, Li Y, Toth JI, Petroski MD, Zhang Z, Zhao JC. N 6-methyladenosine modification destabilizes developmental regulators in embryonic stem cells. Nat Cell Biol 2014; 16 (02) 191-198
- 62 Geula S, Moshitch-Moshkovitz S, Dominissini D. et al. Stem cells. m6A mRNA methylation facilitates resolution of naïve pluripotency toward differentiation. Science 2015; 347 (6225): 1002-1006
- 63 Wang Y, Li Y, Yue M. et al. N 6-methyladenosine RNA modification regulates embryonic neural stem cell self-renewal through histone modifications. Nat Neurosci 2018; 21 (02) 195-206
- 64 Chen Y, Wang J, Xu D. et al. m6A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy 2021; 17 (02) 457-475
- 65 Vu LP, Pickering BF, Cheng Y. et al. The N 6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat Med 2017; 23 (11) 1369-1376
- 66 Yoon KJ, Ringeling FR, Vissers C. et al. Temporal control of mammalian cortical neurogenesis by m6A methylation. Cell 2017; 171 (04) 877-889.e17
- 67 Xu H, Dzhashiashvili Y, Shah A. et al. m6A mRNA methylation is essential for oligodendrocyte maturation and CNS myelination. Neuron 2020; 105 (02) 293.e5-309.e5
- 68 Livneh I, Moshitch-Moshkovitz S, Amariglio N, Rechavi G, Dominissini D. The m6A epitranscriptome: transcriptome plasticity in brain development and function. Nat Rev Neurosci 2020; 21 (01) 36-51
- 69 Koreman E, Sun X, Lu QR. Chromatin remodeling and epigenetic regulation of oligodendrocyte myelination and myelin repair. Mol Cell Neurosci 2018; 87: 18-26
- 70 Sevgi M, Rigoux L, Kühn AB. et al. An obesity-predisposing variant of the FTO gene regulates D2R-dependent reward learning. J Neurosci 2015; 35 (36) 12584-12592
- 71 Li L, Zang L, Zhang F. et al. Fat mass and obesity-associated (FTO) protein regulates adult neurogenesis. Hum Mol Genet 2017; 26 (13) 2398-2411
- 72 Du T, Rao S, Wu L. et al. An association study of the m6A genes with major depressive disorder in Chinese Han population. J Affect Disord 2015; 183: 279-286
- 73 Sun L, Ma L, Zhang H. et al. Fto deficiency reduces anxiety- and depression-like behaviors in mice via alterations in gut microbiota. Theranostics 2019; 9 (03) 721-733
- 74 Li H, Ren Y, Mao K. et al. FTO is involved in Alzheimer's disease by targeting TSC1-mTOR-Tau signaling. Biochem Biophys Res Commun 2018; 498 (01) 234-239
- 75 Ma S, Chen C, Ji X. et al. The interplay between m6A RNA methylation and noncoding RNA in cancer. J Hematol Oncol 2019; 12 (01) 121
- 76 He L, Li H, Wu A, Peng Y, Shu G, Yin G. Functions of N 6-methyladenosine and its role in cancer. Mol Cancer 2019; 18 (01) 176
- 77 Cui Q, Shi H, Ye P. et al. m6A RNA methylation regulates the self-renewal and tumorigenesis of glioblastoma stem cells. Cell Rep 2017; 18 (11) 2622-2634
- 78 Visvanathan A, Patil V, Arora A. et al. Essential role of METTL3-mediated m6A modification in glioma stem-like cells maintenance and radioresistance. Oncogene 2018; 37 (04) 522-533
- 79 Visvanathan A, Patil V, Abdulla S, Hoheisel JD, Somasundaram K. N 6-methyladenosine landscape of glioma stem-like cells: METTL3 is essential for the expression of actively transcribed genes and sustenance of the oncogenic signaling. Genes (Basel) 2019; 10 (02) 141
- 80 Chen M, Wei L, Law CT. et al. RNA N 6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018; 67 (06) 2254-2270
- 81 Han J, Wang JZ, Yang X. et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol Cancer 2019; 18 (01) 110
- 82 Wang Q, Chen C, Ding Q. et al. METTL3-mediated m6A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut 2020; 69 (07) 1193-1205
- 83 Lin S, Liu J, Jiang W. et al. METTL3 promotes the proliferation and mobility of gastric cancer cells. Open Med (Wars) 2019; 14: 25-31
- 84 Cai X, Wang X, Cao C. et al. HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tumor suppressor let-7g. Cancer Lett 2018; 415: 11-19
- 85 Li T, Hu PS, Zuo Z. et al. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer 2019; 18 (01) 112
- 86 Zhu W, Si Y, Xu J. et al. Methyltransferase like 3 promotes colorectal cancer proliferation by stabilizing CCNE1 mRNA in an m6A-dependent manner. J Cell Mol Med 2020; 24 (06) 3521-3533
- 87 Cai J, Yang F, Zhan H. et al. RNA m6A methyltransferase METTL3 promotes the growth of prostate cancer by regulating hedgehog pathway. OncoTargets Ther 2019; 12: 9143-9152
- 88 Cui X, Wang Z, Li J. et al. Cross talk between RNA N 6-methyladenosine methyltransferase-like 3 and miR-186 regulates hepatoblastoma progression through Wnt/β-catenin signalling pathway. Cell Prolif 2020; 53 (03) e12768
- 89 Taketo K, Konno M, Asai A. et al. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int J Oncol 2018; 52 (02) 621-629
- 90 Luo G, Xu W, Zhao Y. et al. RNA m6 A methylation regulates uveal melanoma cell proliferation, migration, and invasion by targeting c-Met. J Cell Physiol 2020; 235 (10) 7107-7119
- 91 Lang F, Singh RK, Pei Y, Zhang S, Sun K, Robertson ES. EBV epitranscriptome reprogramming by METTL14 is critical for viral-associated tumorigenesis. PLoS Pathog 2019; 15 (06) e1007796
- 92 Shi Y, Fan S, Wu M. et al. YTHDF1 links hypoxia adaptation and non-small cell lung cancer progression. Nat Commun 2019; 10 (01) 4892
- 93 Bai Y, Yang C, Wu R. et al. YTHDF1 regulates tumorigenicity and cancer stem cell-like activity in human colorectal carcinoma. Front Oncol 2019; 9: 332
- 94 Zhao X, Chen Y, Mao Q. et al. Overexpression of YTHDF1 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Biomark 2018; 21 (04) 859-868
- 95 Liu T, Wei Q, Jin J. et al. The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation. Nucleic Acids Res 2020; 48 (07) 3816-3831
- 96 Nishizawa Y, Konno M, Asai A. et al. Oncogene c-Myc promotes epitranscriptome m6A reader YTHDF1 expression in colorectal cancer. Oncotarget 2017; 9 (07) 7476-7486
- 97 Lin X, Chai G, Wu Y. et al. RNA m6A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail. Nat Commun 2019; 10 (01) 2065
- 98 Kim DJ, Iwasaki A. YTHDF1 control of dendritic cell cross-priming as a possible target of cancer immunotherapy. Biochemistry 2019; 58 (15) 1945-1946
- 99 Tanabe A, Tanikawa K, Tsunetomi M. et al. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett 2016; 376 (01) 34-42
- 100 Yang N, Ying P, Tian J. et al. Genetic variants in m6A modification genes are associated with esophageal squamous-cell carcinoma in the Chinese population. Carcinogenesis 2020; 41 (06) 761-768
- 101 Fanale D, Iovanna JL, Calvo EL. et al. Germline copy number variation in the YTHDC2 gene: does it have a role in finding a novel potential molecular target involved in pancreatic adenocarcinoma susceptibility?. Expert Opin Ther Targets 2014; 18 (08) 841-850
- 102 Huang H, Weng H, Sun W. et al. Recognition of RNA N 6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol 2018; 20 (03) 285-295
- 103 Gutschner T, Hämmerle M, Pazaitis N. et al. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) is an important protumorigenic factor in hepatocellular carcinoma. Hepatology 2014; 59 (05) 1900-1911
- 104 Müller S, Glaß M, Singh AK. et al. IGF2BP1 promotes SRF-dependent transcription in cancer in a m6A- and miRNA-dependent manner. Nucleic Acids Res 2019; 47 (01) 375-390
- 105 Hämmerle M, Gutschner T, Uckelmann H. et al. Posttranscriptional destabilization of the liver-specific long noncoding RNA HULC by the IGF2 mRNA-binding protein 1 (IGF2BP1). Hepatology 2013; 58 (05) 1703-1712
- 106 Nguyen LH, Robinton DA, Seligson MT. et al. Lin28b is sufficient to drive liver cancer and necessary for its maintenance in murine models. Cancer Cell 2014; 26 (02) 248-261
- 107 Dai N, Zhao L, Wrighting D. et al. IGF2BP2/IMP2-deficient mice resist obesity through enhanced translation of Ucp1 mRNA and other mRNAs encoding mitochondrial proteins. Cell Metab 2015; 21 (04) 609-621
- 108 Simon Y, Kessler SM, Bohle RM, Haybaeck J, Kiemer AK. The insulin-like growth factor 2 (IGF2) mRNA-binding protein p62/IGF2BP2-2 as a promoter of NAFLD and HCC?. Gut 2014; 63 (05) 861-863
- 109 Xu D, Shao W, Jiang Y, Wang X, Liu Y, Liu X. FTO expression is associated with the occurrence of gastric cancer and prognosis. Oncol Rep 2017; 38 (04) 2285-2292
- 110 Deng X, Su R, Stanford S, Chen J. Critical enzymatic functions of FTO in obesity and cancer. Front Endocrinol (Lausanne) 2018; 9: 396
- 111 Chen J, Du B. Novel positioning from obesity to cancer: FTO, an m6A RNA demethylase, regulates tumour progression. J Cancer Res Clin Oncol 2019; 145 (01) 19-29
- 112 Li Y, Zheng D, Wang F, Xu Y, Yu H, Zhang H. Expression of demethylase genes, FTO and ALKBH1, is associated with prognosis of gastric cancer. Dig Dis Sci 2019; 64 (06) 1503-1513
- 113 Tang B, Yang Y, Kang M. et al. m6A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling. Mol Cancer 2020; 19 (01) 3
- 114 Huang Y, Su R, Sheng Y. et al. Small-molecule targeting of oncogenic FTO demethylase in acute myeloid leukemia. Cancer Cell 2019; 35 (04) 677-691.e10
- 115 Yang S, Wei J, Cui YH. et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat Commun 2019; 10 (01) 2782
- 116 Zou D, Dong L, Li C, Yin Z, Rao S, Zhou Q. The m6A eraser FTO facilitates proliferation and migration of human cervical cancer cells. Cancer Cell Int 2019; 19: 321
- 117 Rong ZX, Li Z, He JJ. et al. Downregulation of fat mass and obesity associated (FTO) promotes the progression of intrahepatic cholangiocarcinoma. Front Oncol 2019; 9: 369
- 118 Shriwas O, Priyadarshini M, Samal SK. et al. DDX3 modulates cisplatin resistance in OSCC through ALKBH5-mediated m6A-demethylation of FOXM1 and NANOG. Apoptosis 2020; 25 (3–4): 233-246
- 119 Strick A, von Hagen F, Gundert L. et al. The N 6 -methyladenosine (m6 A) erasers alkylation repair homologue 5 (ALKBH5) and fat mass and obesity-associated protein (FTO) are prognostic biomarkers in patients with clear cell renal carcinoma. BJU Int 2020; 125 (04) 617-624
- 120 Wang T, Hong T, Huang Y. et al. Fluorescein derivatives as bifunctional molecules for the simultaneous inhibiting and labeling of FTO protein. J Am Chem Soc 2015; 137 (43) 13736-13739
- 121 Huang Y, Yan J, Li Q. et al. Meclofenamic acid selectively inhibits FTO demethylation of m6A over ALKBH5. Nucleic Acids Res 2015; 43 (01) 373-384
- 122 Zhuang J, Lin C, Ye J. m6 A RNA methylation regulators contribute to malignant progression in rectal cancer. J Cell Physiol 2020; 235 (09) 6300-6306
- 123 Liu Y, Guo X, Zhao M. et al. Contributions and prognostic values of m6 A RNA methylation regulators in non-small-cell lung cancer. J Cell Physiol 2020; 235 (09) 6043-6057
- 124 Wang J, Zhang C, He W, Gou X. Effect of m6A RNA methylation regulators on malignant progression and prognosis in renal clear cell carcinoma. Front Oncol 2020; 10: 3
- 125 Zuo X, Chen Z, Gao W. et al. M6A-mediated upregulation of LINC00958 increases lipogenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. J Hematol Oncol 2020; 13 (01) 5
- 126 Lu M, Zhang Z, Xue M. et al. N 6-methyladenosine modification enables viral RNA to escape recognition by RNA sensor RIG-I. Nat Microbiol 2020; 5 (04) 584-598
- 127 Chen YG, Kowtoniuk WE, Agarwal I, Shen Y, Liu DR. LC/MS analysis of cellular RNA reveals NAD-linked RNA. Nat Chem Biol 2009; 5 (12) 879-881
- 128 Cahová H, Winz ML, Höfer K, Nübel G, Jäschke A. NAD captureSeq indicates NAD as a bacterial cap for a subset of regulatory RNAs. Nature 2015; 519 (7543): 374-377
- 129 Walters RW, Matheny T, Mizoue LS, Rao BS, Muhlrad D, Parker R. Identification of NAD+ capped mRNAs in Saccharomyces cerevisiae. Proc Natl Acad Sci U S A 2017; 114 (03) 480-485
- 130 Jiao X, Doamekpor SK, Bird JG. et al. 5′ End nicotinamide adenine dinucleotide cap in human cells promotes RNA decay through DXO-mediated deNADding. Cell 2017; 168 (06) 1015-1027.e10
- 131 Zhang H, Zhong H, Zhang S. et al. NAD tagSeq reveals that NAD+-capped RNAs are mostly produced from a large number of protein-coding genes in Arabidopsis . Proc Natl Acad Sci U S A 2019; 116 (24) 12072-12077
- 132 Bird JG, Basu U, Kuster D. et al. Highly efficient 5′ capping of mitochondrial RNA with NAD+ and NADH by yeast and human mitochondrial RNA polymerase. eLife 2018; 7: e42179
- 133 Dumelin CE, Chen Y, Leconte AM, Chen YG, Liu DR. Discovery and biological characterization of geranylated RNA in bacteria. Nat Chem Biol 2012; 8 (11) 913-919
- 134 Miles ZD, McCarty RM, Molnar G, Bandarian V. Discovery of epoxyqueuosine (oQ) reductase reveals parallels between halorespiration and tRNA modification. Proc Natl Acad Sci U S A 2011; 108 (18) 7368-7372
- 135 Li J, Jiang XY, Xu SJ. et al. Medicinal chemistry strategies in seeking coronavirus inhibitors. Yao Xue Xue Bao 2020; 55 (04) 537-553
- 136 Kim D, Lee JY, Yang JS, Kim JW, Kim VN, Chang H. The architecture of SARS-CoV-2 transcriptome. Cell 2020; 181 (04) 914-921.e10
- 137 Zhou H, Gan Y, Li Y, Chen X, Guo Y, Wang R. Degradation of rat sarcoma proteins targeting the post-translational prenyl modifications via cascade azidation/fluorination and click reaction. J Med Chem 2023; 66 (11) 7243-7252
- 138 Gan Y, Li Y, Zhou H, Wang R. Deciphering regulatory proteins of prenylated protein via the FRET technique using nitroso-based ene-ligation and sequential azidation and click reaction. Org Lett 2022; 24 (36) 6625-6630
- 139 Gan Y, Chen X, Li Y, Guo Y, Wang R. Sequential azidation/azolation of prenylated derivatives and a click reaction enable selective labeling and degradation of RAS protein. J Org Chem 2023; 88 (15) 10836-10843
- 140 Li Y, Zhou H, Liu H. et al. Development of nitroso-based probes for labeling and regulation of RAS proteins in cancer cells via sequential ene-ligation and oxime condensation. J Org Chem 2023; 88 (03) 1762-1771
- 141 Wang S, Li Y, Zhou H, Wang L, Wang R. Development of biocompatible ene-ligation enabled by prenyl-based β-caryophyllene with triazoline/selectfluor under physiological conditions. J Org Chem 2022; 87 (13) 8648-8655
- 142 Zhou H, Li Y, Gan Y, Wang R. Total RNA synthesis and its covalent labeling innovation. Top Curr Chem (Cham) 2022; 380 (03) 16
- 143 Gan YF, Li YY, Chen XQ, Guo YY, Wang R. Recent advances in quinone methide chemistry for protein-proximity capturing. Synthesis 2023; 55 (08) 1172-1186
- 144 Wang R, Vangaveti S, Ranganathan SV. et al. Synthesis, base pairing and structure studies of geranylated RNA. Nucleic Acids Res 2016; 44 (13) 6036-6045
- 145 Li Y, Zhou H, Chen S. et al. Bioorthogonal labeling and profiling of N 6-isopentenyladenosine (i6A) modified RNA. Nucleic Acids Res 2024 (e-pub ahead of print).